Ns. sialiinimutaation katsotaan päästäneen valloilleen ihmisen aivojen kasvun. Se mutaatio on poistanut kokonaan erään solujen ulkopintojen kemikaalin, joka on muilla eläimillä aivoissa haitallinen, mutta lihas- ja luukudoksissa takaa niille noin nelinkertaisen lujuuden, ja siten nelinkertaiset voimat, kuin ilman sitä.
Sialiinilla on luuultavasti estävä vaikutus glia-solujen kasvuun.
https://asiakas.kotisivukone.com/files/kansanaani.kotisivukone.com/4-08.pdf
" Jared Diamond ei pitkälle puusta pääse
Maailmalla on eräänlaiseksi muotifilosofiksi noussut yhdysvaltalainen fysiologian ja maantieteen professori Jared Diamond.Hänen työpaikkansa on Los Angelesissa si- jaitseva University of California. Myös Suomen tiedemaailman Diamond on hurman-nut niin, että presidentti Tarja Halonen nimitti hänet muutama vuosi sitten Suomen Akatemiaan ulkomaiseksi akateemikoksi. Hän on kirjoittanut neljä suomennettua kir-jaa:´Miksi seksi on hauskaa´,’Tykit,taudit ja teräs’,’Romahdus’ ja ’Kolmas simpanssi’.

Luin kesän aikana hänen teoksensa ’Kolmas simpanssi’, joka kirjan takakannen mu-kaan tutkii simpanssin ja ihmisen geneettistä eroa:”Vaikka ihmisen geeneistä 98 pro- senttia on samoja kuin simpanssilla, lajimme on kehittynyt täysin ainutlaatuiseksi”.
Jos tutkija nimetään jäseneksi Suomen Akatemiaan olettaisi, että hänellä olisi myös riittäviä ansioita tähän nimitykseen. Kirjan ’Kolmas simpanssi’ antaa ymmärtää, että Diamond saa irti jotain perustavanlaatuista uutta esiin ihmisen kehityksestä apinasta ihmiseksi. Juuri tätä Diamond ei kuitenkaan tee. Hän on tutkinut apinoita ja lintuja etenkin Uudessa Guineassa ja perustanut luonnonsuojelualueita sinne.Hän on myös tutkinut kansojen kieliä ja toteaa,että yksinomaan Uudessa Guineassa on yli tuhat kieltä; kieli voi vaihtua toiseksi jopa 20 kilometrin välein.Jos hänellä joitain tieteellisiä ansioita on, niin ne olisivat juuri luonnon tai kielten tutkijana. Sen sijaan ihmisen kehityksen tutkijana hän ei pääse puusta pitkään.
Diamondin kirja ’Kolmas apina’ sisältää yli 400 sivua, joista alkuosa - melkein puolet koko kirjasta selvittelee ihmisten ja eläinten sukupuolisen käyttäytymisen vertailua. Eikä hän vertaile vain apinoita ja ihmistä, vaan vertailuun tulee myös koko lintumaail- ma. Juuri sitä hän tekee myös kautta ensimmäisen suomennetun kirjansa, joka on täysin sosiobiologistinen, pavlovilaista ehdollistumista sen enempää ihmisillä kuin eläimilläkään tuntematon humpuukiteos. Hän yrittää geenien muunnosten kautta selvittää, kuinka ihminen erosi 6 miljoonaa vuotta sitten simpanssien kehityslinjasta. Lukija saa kyllä tästä sellaisen käsityksen, että geeneissä tapahtunut kehitys johti ihmiseen. Diamondilta jää selvittämättä, miksi näin tapahtui.
Diamondin virhe on, että hän tutkii ihmisen kehitystä pelkästään geenien kautta ja pseudoluonnontieteilijänä poistaa kokonaan sen, mitä ihmisen toiminnan yhteiskun- nallinen luonne kehitykselle on merkinnyt.Tässä hänen pitäisi tutustua Engelsin kah- teen merkittävään kirjoitukseen,Työn osuus apinan kehittymisessä ihmiseksi’,joka on yksi Engelsin viimeisistä kirjoituksista ja julkaistu Die Neue Zeit -lehdessä v.1895-96, sekä ’Perheen, yksityisomaisuuden ja valtion alkuperä’, kirjoitettu välillä maaliskuu - toukokuu 1884. Vaikka nämä Engelsin teokset ovatkin 1800-luvulta ja vaikka kaikki luonnontieteet ovat kehittyneet valtavasti sen jälkeen, näillä Engelsin kirjoituksilla on kuitenkin erittäin perustavanlaatuinen merkitys.
Sitä paitsi Diamond ei tunne ihmisen evoluutiogenetiikankaan TODELLISIA tuloksia, kuten mm. Ajit Varkin osoittamaa sialiinimutaatiota. Uusimpien tutkimusten mukaan suurimman eron ihmisen ja simpanssin keskushermostojen välille aiheuttaa se, että solupintojen sokerit poikkeavat melko jyrkästi lajien välillä. Simpanssin ja muiden ”alempien” nisäkkäiden sialiini on korvautunut ihmisellä suurella joukolla monimuo-toisempia sokeriyhdisteitä.Juuri tämän arvelee Kalifornian San Diegon yliopiston So-keribiologian tutkimuskeskuksen johtaja professori Ajit Varki olevan ihmisen aivokuo- ren simpanssiin nähden suunnattomasti tehokkaamman ehdollistumiskapasteetin taustalla (New Scientist 26.10.2002 Sugar Rush: Did sugars make us smart?).
Kokeet osoittavat, että sellaisilla eläimillä kuin hiirillä ja apinoilla sialiinitaso on aivois-sa paljon matalampi kuin muissa kudoksissa, mutta vain ihmisellä kyseinen aine puuttuu kokonaan, ja se puuttui myös neanderdalin ihmiseltä. (Tämän on osoittanut ruotsinvirolainen Svante Pääbo saksalaisesta Max Planck -instituutista.Ei olisi tuosta ”unohduksesta” seurannut pelkästään hyvää Saksan, Ruotsin tai Viron ”akateemikolle”…)
Toisaalta sialiinimutaatio heikentää lihasvoimaa ja luiden lujuutta jopa neljännekseen entisestä,joten noiden menetysten vastapainoksi siitä on täytynyt olla hyötyä esimer-kiksi oppimiselle. Ja on pitänyt olla yhteiskunta, joka suojaa myös heikompivoimaisia ja löytää näille oikeita tehtäviä, kehityksenkin geneettisenä edellytyksenä, eikä ”jarruna”. Aivan varmasti vastaavanlaisia sialiinimutaatioita on tapahtunut muillakin nisäkkäillä kymmenien vuosimiljardien saatossa, mutta ne ovat karsiutuneet ennen sopivanlaista yhteisöä/yhteiskuntaa!
Luonnontieteellisesti perustava merkitys on I. P. Pavlovin ehdollistumisteorialla (1904, 1927) ja psykologisesti L.S. Vygotskin kielellisellä ajatteluteorialla (jossa kie-len ja ajattelun pohjalla ja veturina on työn ja aineellisten työkalujen kehitys. Jos Dia-mondilla on varaa liittää kirjaansa parisataa lähdeteosta, miksi hänellä on varaa pu-dottaa lähteistään Engels,Pavlov ja Vygostky pois,ja myös jättää ne lukematta? Sen- kö täytistä sellainen tonttu tekee ”Suomen ulkomaisena akateemikkona”, munaa ko-ko maan eikä pelkästään nimittäjäänsä! Kun hän näin tekee, hän on perusteellisesti pihalla koko aihepiiristään.
Engelsin mukaan siis juuri työ loi ihmisen. Laskeuduttuaan puusta alas, ihmisen oli vapautettava kätensä erilaisten aseiden ja työkalujen käyttöön.Se vaati pystykäyntiä, joka taas vaati ennen kuulumatonta aivokapasiteettia sillä menetelmällä, jolla ihmi-nen kävelee kahdella jalalla.Yksittäisenä eläimenä ihminen oli hyvin haavoittuva pe-toja, kuten hyeenoja ja leijonia vastaan,ihminen kykeni puolustautumaan ja saalista-maan vain tiiviinä ryhmänä, se puolestaan vaati kommunikaation kehitystä, josta lopulta seurasi kielen syntyminen.
Tässä yhteydessä voidaan kysyä,mikä merkitys geeneillä oli tässä kehityksessä. To- dennäköisesti ihmisen käyttäytyminen ja geneettiset muutokset tukivat toisiaan:edel- linen muodosti sosiaalisesti periytyvänä vaatimukset jälkimmäiselle työn takia paljon enemmän kuin aikaisemmin. Kun pystykäynnin oli parannuttava,sellaiset geneettiset muutokset pääsivät voitolle,jotka paransivat ihmisen pystykäyntiä.Kun ihmisten tarve viestimiseen tuli tärkeäksi, sellaiset geneettiset muutokset pääsivät voitolle, jotka edistivät ihmisen äänen tuottamista kuten kurkunpään lasku alaspäin. Ja kun tämä kaikki vaati parempia ja suurempia aivoja, geenimuutokset tekivät senkin mahdolliseksi.
Ihmisen kehityksen myötä eteen tuli vaihe, että ihmisen yhteiskunnallinen kehitys ohitti nopeudessaan valtavasti geeneissä tapahtuvan kehityksen. Yhteiskunnallinen kehitys siis lähes poisti geeneihin perustuvan kehityksen merkityksen.Kuitenkin geeneissä tapahtuva kehitys on varmasti vielä olemassa. Geneettinen kehitys on vallinnut maailmassa miljardeja vuosia. Se on olennainen osa elämää. Ei ihmisen parimiljoonainen yhteiskunnallinen/ kulttuurikehitys sitä ole ihmisen osalta kokonaan poistanut. Miten se tulevaisuudessa ilmenee, on hämärän peitossa.
Joitakin hyviä huomioita kirja kuitenkin sisältää - sokea kanakin löytää joskus jyvän. Hänen maantiedettä ja ihmisen kulttuurikehitystä koskevat huomionsa kyllä toimivat. Esim.hän kiinnittää huomionsa Euraasian ja Amerikan erilaisuuteen;Eurooppa ja Aa- sia ovat ”vaakasuorassa”, itä-länsi-suunnassa.Sen sijaan Pohjois- ja Etelä-Amerikka on ”pystysuorassa”, pohjois-etelä-suunnassa. Kaikki kasvilajikkeet levittäytyvät hel- posti samojen leveysasteiden puitteissa idästä länteen ja päinvastoin. Sen sijaan pi- tuusasteiden mukaan etelästä - pohjoiseen ja päinvastoin - levittäytymien vaatii aina uusien lajikkeiden syntymistä.Lisäksi hän toteaa aivan oikein,että suurten vetojuhtien kesyttäminen Euroopan ja Aasian alueilla paransi maanviljelyksen mahdollisuuksia ja siten ihmisen kulttuurikehitystä. Sen sijaan Uudessa Guineassa ei ollut isoja eläimiä kesytettäviksi ja siksi kehitys Uudessa Guineassa jäi Euroopasta jälkeen.
Kun Diamond yrittää selvittää ihmisen puhekielen syntymistä, hänen järkeilynsä jää valovuosien päähän parhaista marxilaisista teorioista. Hän ei pääse käsitteiden syntymiseen ollenkaan.Hän olettaa Noam Chomskya mukaillen,että ainakin kielioppi on ihmisessä geneettisesti määräytynyttä. Hän ei ymmärrä lähteä siitä, että kieli on objektiivisen todellisuuden heijastumaa,ja että jokaisella käsitteellä on lähtökohtansa objektiivisessa todellisuudessa.Ei myöskään kielioppia pidä etsiä geenien kautta, se syntyy luonnostaan ihmisen sosiaalisessa vuorovaikutuksessa.
Diamond perustelee käsityksiään ihmislapsen nopealla kehityksellä. Hän pitää ihmeenä, kuinka 5-vuotias englantilainen lapsi osaa paremmin englantia, kuin moni ei-englantilainen 20 vuoden opiskelun jälkeen. Ihmislapsi syntyykin avuttomana, mutta siitä syystä sen onkin kehityttävä nopeasti.Ei sillä ole mitään tekemistä kieliop- pien kanssa. Vaikka ihminen kehittyykin nopeasti,kehitys vie kuitenkin kauan – vielä parikymppisinä suuri osa ihmisistä on vielä täysin raakileita.
Diamondin kirjan loppuhuipennus viimeistään osoittaa, ettei hän ole päässyt puusta pitkään.Hän erittelee ihmisen peruspiirteiksi = perusluonteeseen kuuluviksi - ihmisen kyvyn käyttää työkaluja, ja ihmisen puhekielen, mitkä varmasti ovatkin ihmisen oleel- lisia tuntomerkkejä. Sen lisäksi hän liittää ihmisen perusluonteeseen taipumuksen huumeiden käyttöön ja taipumuksen massamurhiin.
Marxilaisuus kieltää koko käsityksen ihmisen perusluonteesta ja selittää kaiken käyt- täytymisen ihmisen yhteiskunnallisten vuorovaikutussuhteiden kautta. Ihminen ei ole ihminen, ellei hän ole kasvanut yhteiskunnassa,vaan lähinnä harvinaisen avuton ”monivammainen simpanssi”. Diamondin osalta näyttää siltä, että hän on liittänyt ih-misen perusluonteeseen koko sen amerikkalaisen maailman,mikä häntä ympäröi. Ih-me, ettei hän liitä ihmisen perusluonteeseen kuuluvaksi myös hampurilaisen symistä McDonaldsin pikaruokalassa.Diamondin kirja kannattaa kuitenkin lukea ja siivilöidä siitä käytettävissä oleva aines omaan käyttöön kuten juuri tuo maantieteen vaikutus ihmisen kulttuurikehitykseen.
Reijo Katajaranta "
https://www.tiede.fi/artikkeli/uutiset/jared_diamondista_akateemikko
15.10.2004 klo 0:00
Helsingissä parhaillaan vierailevasta yhdysvaltalaisesta tutkijasta ja tietokirjailijasta Jared Diamondista, 67, tulee seitsemästoista ulkomainen tiedemies, joka kantaa akateemikon arvonimiä. Arvonimi luovutetaan tänään Suomen Akatemiassa pidettävässä ...
Helsingissä parhaillaan vierailevasta yhdysvaltalaisesta tutkijasta ja tietokirjailijasta Jared Diamondista, 67, tulee seitsemästoista ulkomainen tiedemies, joka kantaa akateemikon arvonimiä. Arvonimi luovutetaan tänään Suomen Akatemiassa pidettävässä juhlassa.
Diamond on Kalifornian yliopiston (Los Angeles) maantieteen professori.Hän on kan- sainvälisesti arvostettu tutkija, joka on erityisesti kirjoillaan luonut siltaa luonnontie-teellisen ja humanistisen tutkimuksen välille.Diamond voitti Pulitzer-palkinnon vuon-na 1998 teoksellaan Guns,Germs, and Steel:The Fates of Human Societies.Se on il-mestynyt suomeksi nimellä Tykit,taudit ja teräs.Ihmisen yhteiskuntien kohtalot (Terra Cognita 2003).Suomeksi on kääntynyt myös Miksi seksi on hauskaa? (WSOY 1998)
Tasavallan presidentti Tarja Halonen myöntää 12-jäsenisen Suomen akatemian halli-tuksen esityksestä akateemikon arvonimen erittäin ansioituneelle suomalaiselle tai ulkomaiselle tieteenharjoittajalle.Peräti 28 ulkomaiselle tiedemiehelle on tähän men-nessä annettu akateemikon arvo. Arvonimi voi olla samanaikaisesti enintään kahdel-latoista kotimaisella tieteenharjoittajalla. Ulkomaisten akateemikkojen määrää ei sen sijaan ole rajoitettu.
***
Hankitut ominaisuudet syrjäyttävät evoluutiossa geneettisiä immuniteetissa
www.newscientist.com/article/dn21876-wa ... agues.html
" Was humanity born in the mother of all plagues?
* 20:01 04 June 2012 by Michael Marshall
Around 100,000 years ago, the human race was on the brink of extinction. Confined to Africa, our population had fallen to less than 10,000. Yet within a few tens of thousands of years, we began spreading around the world.
New genetic evidence suggests that one factor contributing to the population bottle- neck was a massive epidemic of bacterial disease. The bacteria were exploiting two immune system genes, turning them against us. So the solution was simple: get rid of the traitorous genes.
Ajit Varki of the University of California, San Diego and colleagues looked at two genes called Siglec-13 and Siglec-17. Both code for proteins that are involved in controlling the immune system, helping to decide whether immune cells should go on the offensive.
Varki found that both genes are active in chimpanzees,but not in humans.Siglec-13 has been entirely deleted from the human genome,while Siglec-17 is non-functional as a result of losing one letter from its code.
Traitor genes
Why would we have got rid of two useful immune genes? Varki reconstructed the lost proteins and found that two dangerous bacteria, Group B Streptococcus and Escherichia coli K1, could bind to them.
Wondering if the bacteria could exploit the proteins, he expressed each protein in some human immune cells. The modified cells had a weaker response to the bacte- ria than immune cells without the proteins. That suggests the bacteria had found a way to dampen the immune response by binding to the two proteins.
Varki thinks that early humans were confronted with a massive epidemic of bacterial infection. The two bacteria he studied are particularly dangerous to newborn babies, who often die after being infected. That could explain why the human population fell so precipitously, and why we got rid of the Siglec genes that made us so vulnerable.
Population crash
The genetic data suggests that the two genes were switched off in some humans between 440000 and 270000 years ago,before modern humans split from our Neanderthal and Denisovan cousins. But it took a long time for the effect to spread through the entire population: some people may have had working versions of Siglec-13 as recently as 46,000 years ago. During that long period, Varki thinks our ancestors were decimated by disease.
"The recent advances in ancient DNA studies and the human genome project have made it possible to look at the co-evolution of humans and pathogens," says Isabelle de Groote of the Natural History Museum in London,UK, who was not involved in the study. By combining data from genetics, archaeology and other disciplines, we can build up a more detailed picture of our evolution.
Journal reference: Proceedings of the National Academy of Sciences, DOI:10.1073/pnas.1119459109 "
Tähän sisältyy järisyttävä evoluutiobiologinen tietääkseni ensi kertaa todistettu ilmiö geenideterminististen ja oppivien järjestelmien vuorovaikutuksesta evoluutiossa, että taudinaiheuttajat voivat evoluoitua geneettisesti käyttämään hyväkseen immunigeenejä sen sijaan että nuo menetyksellisesti torjuisivat ne.
Tämä kannattaa muistaa myös käyttäytymismuotojen evoluution tutkimuksessa: eh-dollistuneet reagointitavat pystyvät käyttämään hyväkseen vihollis- ja saaliseläinten ehdottomia refleksejä. Ehdollistumismekanismi on suorastaan räätälöitynyt siihen, kun se erottaa toiminassa kaiken lainalaisesti toistuvan satunnaisesta ja ainutkertaisesta. "
Sokerit mieltä muovaamassa
- Sokeriko erottaa meidät apinoista
Pakanasanomienkin palstoilla on käyty keskustelua ns. tieteellisen ihmiskuvan kysy-myksistä.Keskustelu on koskenut ihmisen ajattelun, tietoisuuden, tahdon ja psyyk-kisten toimintojen suhdetta lähimpien eläinsukulaistemme psyyken ominaisuuksiin. Edellä mainittuja kysymyksiä voidaan tutkia tieteellisesti ja yksi tällaisista tieteena-loista on v. 1904 lääketieteen nobelistin I. P. Pavlovin aikanaan perustama neurofysiologia, jonka piirissä kehitys on viime aikoina ollut erityisen nopeaa.
Uusimpien tutkimusten mukaan suurimman eron ihmisen ja simpanssin keskusher-mostojen välille aiheuttaa se,että solupintojen sokerit poikkeavat melko jyrkästi lajien välillä. Simpanssin sialiini (siaalihappo, sialic acid) on korvautunut ihmisellä suurella joukolla monimuotoisempia sokeriyhdisteitä. Juuri tämän arvelee Kalifornian San Diegon yliopiston Sokeribiologian tutkimuskeskuksen johtaja professori Ajit Varki olevan ihmisen aivokuoren simpanssiin nähden suunnattomasti tehokkaamman ehdollistumiskapasiteetin taustalla
(New Scientist 26.10.2002 Sugar Rush: Did sugars make us smart?).
https://www.newscientist.com/article/mg17623665-100-sugar-rush/
Kokeet osoittavat, että sellaisilla eläimillä kuten hiirillä ja apinoilla sialiinitaso on ai-voissa paljon matalampi kuin muissa kudoksissa, mutta vain ihmisellä kyseinen aine puuttuu kokonaan, ja se puuttui myös neanderdalin ihmiseltä. Varki ja muut valmiste-levat kokeita, joissa hiiren aivoista poistetaan sialiini, ja katsotaan mitä ilmenee.
Sokerit toimivat erilaisina "kynsinä", "kytkiminä" ja "tunnistusnappeina" kaikkien solujen pinnalla, mutta aivokuorella niillä näyttäisi olevan aivan erityinen rooli juuri ehdollistumisjärjestelmässä. Niillä on nimittäin geenien koodaamiin valkuaisaineisiin nähden suunnattomasti suurempi monimuotoisuus ja sitä tietä potentiaalinen informaatiokapasiteetti.Professori Ajit Varki kirjoittaa mainitussa lehdessä: "Jos kysytte, mikä on tietyn solutyypin glykomi (termi on analoginen genomille, eli solun kaikkien geenien, ja proteomille, sen kaikkien proteiinityyppien joukolle), niin se on monituhatkertaisesti sen genomia monimutkaisempi".
Edelleen kirjoituksessa todetaan,että tuo "monituhatkertaisesti" antaa kovasti aliarvi- oidun kuvan todellisesta tilanteesta.Sokerit koostuvat noin kymmenestä "alkeispalas- ta", kun geenit koostuvat kahdesta (adeniini-tymiini ja sytosiini-guamiini, jotka lisäksi voivat olla kahdella tavalla ketjussa). Kaksi noista kymmenestä on jokaiselle tutut glukoosi (rypälesokeri) ja fruktoosi (hedelmäsokeri, ja niiden yhdistelmä on sitten tavallinen ruokosokeri). Nuo palat voivat kuitenkin liittyä toisiinsa monin eri tavoin, esimerkiksi erilaisissa kulmissa. Kuvaavaa on, että kun kuuden askelman DNA-pätkällä on 4^6 = 4096 mahdollista muotoa, niin kuuden alkeissokerirenkaan glykosaminoglykaanilla mahdollisia muotoja on 12 miljardia!
Solussa monimutkaisia sokereita syntetisoi Golgin laite entsyymien avulla, jotka ovat geenien määräämiä. Geenit eivät siis suoraan koodaa sokereita, ja välillisestikin ne voivat periaatteessakaan määrätä eksaktisti vain joitakin tärkeimpiä yksinkertaisia sokereita. Lopullisen muotonsa monimutkaiset molekyylit saavat ympäristö- ja solunulkoisistakin syistä erityisesti solujen pinnalla.
San Diegon tutkimukset antavat mahdollisuuden olettaa, että monimutkaisilla sokeri-molekyyleillä eli hiilihydraateilla, joita tähän asti on pidetty enemmänkin solun ener-giavarastona, on merkitystä ihmisen psyykkisten prosessien muodostajana. Jos tule-vaisuuden tutkimukset vahvistavat tämän, se tulee olemaan tärkeä askel sekä tajun-nallisten ilmiöiden demystifioimisen että niiden sellaisen vääränlaisen liian suoravii-vaisen biologisoimisen torjumiseksi, jonka mukaan kaikki informaatio aivoissamme olisi vahvasti geenien määräämää.
Keskushermostomme tai geenistömme "informaatiokapasiteetti" ei siinä tapaukses-sa aseta mitään esteitä psyykkisten prosessien materialistisen perustan suunnalta niiden tieteelliseksi selittämiseksi. Lupaavalta vaikuttaa silloin Darwinin, Pavlovin, ns. kielellisen ajatteluteorian (mm. Vygotsky), ja viimeisimpänä lenkkinä University of California San Diegon tutkijoiden mm. Ajit Varkin ja Marty Serenon tutkimuslinja.
***
"Mutaatio, joka pelasti meidät malarialta auttaa antropologeja"
https://22century.ru/biology-and-biotechnology/55998
Мутация, которая спасла нас от малярии, теперь поможет антропологам

Австралопитек анамский, чьи останки найдены в Аллия Бей, Кения. Реконструкция выполнена Олегом Осиповым для АНТРОПОГЕНЕЗ.РУ.
Australopithecus anamensis, ihmisapina, jonka jäänteitä on löydetty Kenian Allia Bayssa (Meave Leakey). Ei sialiimutaatiota.
Мы все воодушевлены успехами палеогенетики на поприще изучения эволю-ции человека. Правда, есть одно НО: подавляющее большинство генетических исследований касаются ископаемых материалов за пределами Африки. А как же прародина человека? Земли,нашпигованные костями наших предков - Кени-я, Танзания, ЮАР? Костей много,а с ДНК всё плохо: тёплые и влажные условия губительны для нуклеотидных цепочек.Самый древний африканец, геном кото-рого удалось «пощупать» - юноша из Баллито Бэй,чей возраст всего-то 2 тыс. лет.Что же делать исследователям Чёрного континента? Искать обходные пути.
Afrikassa tutkimuksen ongelmana on, että DNA ei säily sikäläisissä olosuhteissa. Ballito Bayn poika n. 2000 vuotta sitten Etelä-Afrikan länsirannikolta on aikaisin afrikkalainen, josta on saatu talteen DNA.Hän kuului khoisan- eli hottentottiväestöön. On etsittävä muita tutkimusmenetelmiä.
Помимо ДНК, в костях могли уцелеть другие органические соединения, из которых надо постараться извлечь максимум информации. Мы уже писали о протеомном aнализе — исследовании аминокислотного состава коллагена, сохранившегося в костях древних людей.
DNA:n ohella luissa voi säilyä muita orgaanisia yhdisteitä, joista pitää yrittää saada irti maksimaalisesti informaatiota. Kirjoitimme jo proteomianalyysista, kollageenin aminohappokoostumuksen analyysistä, jota on säilynyt muinaisten ihmisten luissa.
В статье, опубликованной в PNAS,предлагается метод, который пока не введён в практику, но сулит перспективы в будущем. Так считает коллектив авторов, среди которых - Мив Лики (Meave Leakey),жена знаменитого антрополога Ричарда Лики (Richard Leakey). В чём суть предлагаемого подхода?
PNASissa julkaistussa artikkelissa esitetään menetelmä, jota ei ole vielä sovellettu käytäntöön, mutta joka tarjoaa lupaavia näköaloja tulevaisuudessa. Näin katsoo tekijäkollektiivi, johon kuuluu myös Richard Leakeyn vaimo Meave Leakey. Mikä on esitetyn lähestymistavan ydin?

Antropologi Meave Leakey.
В центре внимания учёных - определённый тип углеводов, называемый сиало- выми кислотами (Sialic acids). Эти сложные органические соединения входят в состав биополимеров клеток животных и во многом определяют свойства клеточной поверхности.
Tieteilijöiden huomion keskiössä on tietty hiilivetyjen ryhmä jota sanotaan sialiini-hapoiksi (Sialic acids, Sialinsäuren). Nämä monimutkaiset orgaaniset yhdisteet kuuluvat eläinsolujen biopolymeereihin ja monin tavoin määrittelevät solujen pinnan ominaisuudet.
В исследовании речь идёт об одной из сиаловых кислот - Neu5Gc (труднопро-износимое название: N-гликолилнейраминовая кислота). Чем она интересна? Тем, что синтезируется в организме млекопитающих… кроме человека.
Tutkimuksessa on kyse yhdestä siaalihaposta: Neu5Gc (N-glykolyyli-neuramiinihap-po). Mikä siinä on merkittävää? Se, että sitä muodostuu imettäväisten elimistössä - IHMISTÄ LUKUUN OTTAMATTA.
Ген CMAH, отвечающий за синтез Neu5Gc, сломался у какого-то нашего предка 2 — 3 млн лет назад.
Geeni CMAH, joka vastaa Neu5Gc:n synteesistä, rikkoontuui jollakulla esi-isällämme 2 - 3 miljoonaa vuotta sitten.
Почему эта мутация распространилась у людей? Потому, что сиаловой кисло-той Neu5Gc пользовались различные патогены, в частности, малярийный плаз-модий Plasmodium reichenowi, для вторжения в клетку (именно поэтому этот патоген вызывает малярию у шимпанзе, но неопасен для человека). Неработа-ющий вариант гена CMAH защищал от малярии,и поэтому распространился в популяции.
Miksi tämä mutaatio levisi ihmisillä?Siksi, että sialiinihappoa Neu5Gc hyödynsivät erilaiset patogeenit, erityisesti malariaeliö Plasmodium reichenowi tunkeutuakseen soluun (juuri siksi tämä patogeeni aihauttaa malarian simpanssilla,mutta on ihmiselle vaaraton). CHMAN-geenin toimimaton variantti suojaa malarialta, ja siksi se yleistyi populaatiossa.

The Plasmodium parasite has evolved over time, losing and gaining genes involved in invasion of red blood cells. This suggests that the recognition of red blood cells from new species may be one of the key barriers that parasites need to overcome in order to infect different primate hosts.
Исследования показали, что мутация вдобавок создала репродуктивный барь-ер: иммунная система носителя инактивированного гена распознавала сперма-тозоиды, содержащие Neu5Gc, как чужие, и отторгала их. Популяция должна была разделиться на две нескрещивающиеся, состоящие из Neu5Gc-положительных и Neu5Gc-отрицательных особей.
Tutkimukset osoittivat, että (sialiini)mutaatio muodosti lisäksi lisääntymisesteen: inaktivoituneen mutatoituneen geenin kantajan immuunijärjestelmä torjui "vieraina" siittiöt, jotka sisälsivät Neu5Gc:ä, eläinten sialiinia. Populaatio jakautui kahdeksi keskenään risteytymättömäksi: sialiinipositiiviseksi ja sialiininegatiiviseksi.
Последние — прямые предки современных людей.
Jälkimmäiset olivat meidän esi-isiämme.
На самом деле Neu5Gc встречается в крови и тканях человека, но в очень маленьких («следовых») количествах. Видимо, она попадает в наш организм с пищей, прежде всего с красным мясом. Исследователи провели эксперимент: вывели генетически модифицированных мышей с инактивированным геном CMAH, давали им разные продукты и наблюдали за составом крови. Неболь-шой процент Neu5Gc встречался в крови только тех грызунов, которых пичкали содержащим этот углевод кормом.
Itse asiassa sialiinia Neu5Gc tavataan myös ihmisen veressä ja kudoksissa mutta erittäin pieninä määrinä (jääminä). Nähtävästi sitä päätyy elimistöömme ruoista, erityisesti punaisesta lihasta. Tutkijat suorittivat kokeen: tuottivat geennimuunellun hiirikannan inaktivoidulla sialiinin CMAN-geenillä, syöttivät niille erilaisia ruokia ja tarkkailivat veren koostumusta. Hyvin pieni määrä Neu5Gc:ä osoitettiin vain niiden jyrsijöiden verestä, joita oli ruokittu tätä hiilivetyä sisältävällä rehulla.
Итак, если бы мы смогли найти Neu5Gc в костях древних гоминид, у нас появи- лся бы способ отличить наших прямых предков от их тупиковых родственников. А небольшой процент Neu5Gc в останках очевидных Homo указывал бы на то, что эти люди уже потребляли много мяса.Оставалось найти способ,как уловить в ископаемых эту самую сиаловую кислоту.
Noin ollen, jos voisimme löytää sialiinia Neu5Gc muinaistan hominidien luista, oli-simme saaneet keinon erottaa esi-isämme umpikujakehityslinjojen yksilöistä.Ja pieni määrä sialiinia ilmeisten Momo-yksilöiden luissa tarkoittaisi, että nämä söivät yhä paljon lihaa. Tehtäväksi asettuu löytää tapa, miten eristää luujäänteistä tätä samaa sialiinhappoa.
Эврика! Исследователи нашли такой способ.Оказывается,в результате распада Neu5Gc его производная часть включается в состав хондроитинсульфата - од-ного из компонентов костной ткани.Такой вариант хондроитинсульфата обозна- чается Gc-CS. Можно ли надеяться, что он сохранился в ископаемых костях?
Heureka! Tutkijat löysivät sellaisen menetelmän. osoittautuu,että Neu5Gc:n hajoami- sen tuloksena sen hajoamistuote kytkeytyy kondroitiinisul(faa)tin - erään luukudok-sen ainesosan koostumukseen. Sellaisesta kondroitiinisulfaatin variantista käytetään merkintää Gc-CS. Olisiko sellainen saattanut säilyä luulöydöissä?
Для проверки исследователи взяли относительно «молодые» останки животных - пещерного медведя, мамонта и лося, возрастом от 12 до 50 тыс. лет. 100 мг образца оказалось достаточно, чтобы обнаружить Gc-CS. Окрылённые успехом учёные подвергли анализу 100 мг кости Homo erectus, жившего на Яве более миллиона лет назад. Увы! Хондроитинсульфат в образце нашёлся, но в количестве, недостаточном для анализа.
Todisteeksi tukijat ottivat suhteellisen "nuoria" eläinten jäänteitä: luolakarhun, mam-mutin, hirven, iältään 12000 - 50000 vuotta. 100:n mg:n koepala osoittatui riittäväksi Gc-CS:n osoittamiseksi. Menestyksen innoittamat tutkijat ottivat analysoitavakseen 100 mg Homo erectuksen luuta, joka oli elänyt Jaavalla yli miljoona vuotta sitten. Kondroitiinisulfaattia näytteessä oli, mutta liian vähän analyysia varten.
Не успокоившись, учёные пошли дальше. Они проанализировали останки жи-вотных возрастом около 4 млн лет (из местонахождения Аллия Бей на восточ-ном берегу озера Туркана в Кении, где Мив Лики нашла анамского австралопи-тека). Только на этот раз взяли кусочки костей побольше - пятиграммовые. К радости исследователей, на этот раз результат оказался положительным.
Tutkijat etenivät väsymättömästi. He analysoivat noin 4 miljoonan vuoden ikäisiä eläinten jäänteitä (Alley Bayn alueella Keniassa Turkana-järven itärannalla, josta Meave Leakey löysi Australopithecus anamnensiksen). Tällä kertaa kudosta otettiin enemmän, noin 5 grammaa. Tutkijoiden iloksi tämä koe osoittatui positiiviseksi (siis ELÄIMEN sialiinia).
Что это нам даёт? Раз Gc-CS сохраняется в 4-миллионолетних останках, а инактивация гена CMAH случилась 2-3 млн лет назад, то, анализируя останки австралопитеков и ранних Homo, можно попытаться поймать этот переломный момент эволюции. Ископаемых гоминид можно будет разделить Gc-CS-положи-тельных и Gc-CS-отрицательных, тем самым отделяя ветвь, ведущую к совре-менному человеку,от вымерших неудачников.Появление «следовых» количеств Gc-CS в останках позволит отличить мясоедов от вегетарианцев.
Mitä tämä meille kertoo? Koska Gc-CS on jäljellä 4 miljoonan vuoden takaisissa jäänteissä, mutta inaktivoitu sialiinigeeni on löydettävissä 2 - 3 miljoonan vuoden ikäisissä jäänteissä, niin Australopithecus- ja Homo-lajeja analysoimalla on mahdol-lista paikallistaa tämä kehityksen "saranahetki". Löydetyt hominidit on jaettava Gc-CS-negatiivisiin ja Gc-CS-positiivisiin, samalla jaken haaraan, joka on johtanut nyky-ihmiseen ja niihin toisiin, sukupuuttoon kuolleisiin "huono-onnisiin". Hyvin pienet jäämä-sialiinin määrät auttavat tekemään jaon lihansyöjiin ja kasvissyöjiin.
Какова последовательность событий? Сначала наши предки перестали синте-зировать Neu5Gc, потом стали есть мясо,или наоборот? Ответ должны дать ис- следования африканских костей.Особенно интересно применить новый подход к спорным ископаемым — типа австралопитека седибы или Homo naledi из Юж-ной Африки.Кроме того,хочется протестировать азиатских древних Homo, что-бы понять:когда их предки покинули Африку? До роковой мутации (то есть до 2 млн лет назад) или после? Всё это можно будет сделать…в будущем, когда ме-тод усовершенствуют. Пока что требуемый образец - 5 граммов - слишком ве-лик, и это затрудняет применение методики к редким и ценным останкам древ-них гоминид.Что ж,авторы надеются,что им удастся довести методику до совер- шенства,и в скором будущем протестировать на каких-нибудь ископаемых люд- ях,оставивших нам много костей.Например,подойдут Homo georgicus из Дмани- си. Или упомянутые наледи. На полторы тысячи костей 5 граммов для анализа точно найдётся.
Millainen on ollut tapausten kulku? Lakkasivatko esi-isämme ensin syntetisoimasta sialiinia Neu5Gc ja sitten alkoivat syödä lihaa vai päinvastoin? Vastata voi afrikka-laisten luiden tutkimus.Erityisen mielenkiintoista on uuden menetelmän soveltaminen uusiin löytöihin tyyppiä Australopithecus sediba ja Homo naledi Etelä-Afrikasta. Näi-den lisäksi on testattava muinaiset aasialaiset Homot saadaksemme tietää, milloin esi-isämme lähtivät Afrikasta.Ennenkö kohtalokasta mutaatiota (eli yli 2.5 miljoonaa vuotta sitten) vai sen jälkeen? Kaikki tämä voidaan tehdä tulevaisuudessa - kun me-netelmä kehittyy. Toistaiseksi vaadittu koepala, 5 grammaa,on liian suuri,ja tämä vai- keuttaa käyttämästä menetelmää muinaisten hominidien harvinaisiin ja arvokkaisiin jäännöksiin. Tekijät toivovat, että heidän onnistuu täydellistää menetelmä ja lähitule-vaisuudessa testata läpi joitakin luulöytyjä,joissa on paljon luita. Esimerkiksi käyvät Homo georgicus Dmanisista. Tai mainitut naledit. Puolestatoita tuhannesta luusta 5 grammaa varmaan joutaa analysoitavaksi.
Myös Denisovan ihmisellä oli sialiinimutaatio
ДНК, подвинься! Древний белок помогает там, где бессильна палеогенетика

Видовую принадлежность останков древнего человека установили по содержащемуся в них коллагену.
Недавно я писал о челюсти денисовского человека, найденной в тибетской пе-щере Байшия. Находка уникальная: наконец-то денисовца обнаружили за пре-делами Денисовой пещеры. Но это исследование замечательно ещё по одной причине. Впервые вид древнего человека удалось идентифицировать по соста-ву белка, сохранившегося в его зубах. Владелец челюсти умер 160 тыс. лет на-зад, его плоть истлела,ДНК распалась на крошечные неопределимые фрагмен- ты, но в глубине зубов сохранился,пусть сильно попорченный временем, но всё ещё различимый для исследователей белок коллаген.В его составе исследова- тели нашли одну аминокислоту,которой нет ни у современного человека, ни у неандертальцев - последовательность совпала только с денисовским челове-ком (у исследователей не было коллагена из Денисовой пещеры, но если есть ДНК, из неё можно получить аминокислотный состав белка).
Справедливости ради надо сказать,что подобные исследования останков древ- них людей делались и раньше. В 2016 году с помощью протеомного анализа идентифицировали фрагментарные человеческие останки,найденные во фран- цузском гроте Оленя. Удалось не только установить, что это неандерталец, но выяснить, что кости принадлежали грудному младенцу — традиционный ДНК-анализ не позволил бы узнать такие де-тали. Но помимо состава коллагена, учёным в тот раз помог анализ митохондриальной ДНК. В случае денисовца с Тибета,«личность» установили только путём исследования древнего коллагена.
Теперь в журнале Nature вышла статья, посвящённая преимуществам новой методики, залихватски названная «Подвинься,ДНК!» Чем же плох уже привыч- ный нам анализ древних генетических последовательностей? Бесспорно, палеогенетика произвела революцию в исследованиях ископаемых останков.
И всё же возможности метода не бесконечны. Во-первых, время не щадит ДНК - через несколько сотен тысяч лет даже в самых лучших условиях от генома не остаётся ничего.Поэтому собранные учёными генетические данные, после мно- гих лет изысканий, касаются, фактически, только 3 видов - неан-дертальцев, денисовцев и древних сапиенсов, и редко выходят за пределы последних 100 - 150 тыс. лет. Исключение - исследование останков из пещеры Сима де лос Уэ-сос в Испании, возраст которых 430 тыс. лет (авторы статьи относят этих гоми-нин к ранним неандертальцам). А ведь так хочется добраться до более древних находок! Выяснить, кто из древних обитателей Евразии ближе всего к общему предку неандертальцев и денисовцев.
Уточнить родственные связи азиатских и европейских гоминин. Разобраться на-конец с гейдельбергским человеком: он предок только неандертальцев или са-пиенсов тоже? (Авторы статьи указывают хронологический интервал жизни ви-да Homo heidelbergensis: 700-200 тыс. лет. А «ранние неандертальцы» из Симы Уэсос - это что-то принципиально другое? Неувязочка).
А ещё дальше в прошлое? Перешагнуть за миллион лет — там ещё больше вопросов. Родословная Homo erectus. Корни и судьба африканских и азиатских форм. Добраться бы до самых ранних Homo. Эх, мечты, мечты…
Есть и вторая проблема: сохранность ДНК сильно зависит от условий, не в по-следнюю очередь - от температуры. 100-тысячелетний образец из Сибири всё ещё хранит достаточно генетического материала,в то время как в 10 раз более молодых останках из Африки уже ничего не осталось.Потому так и не удалось палеогенетикам ничего извлечь даже из таких относительно «юных» находок, как скелет хоббита с острова Флорес.
Коллаген гораздо более стабилен, чем ДНК, и потому для его анализа годится гораздо более широкий спектр палеонтологических находок - и более южных, и более древних. А значит — открывается реальная перспектива найти ответы на перечисленные выше вопросы, включая извечный: кто был прямым предком сапиенса?
Конечно, идея анализа древних белков не нова.Ещё в середине XX века прово- дились эксперименты по поиску аминокислот в ископаемых костях. Но выявить отдельные аминокислоты мало, из них нужно реконструировать состав конкрет- ных белков. Один из разработчиков методики, биоархеолог из Копенгагена Мэтью Коллинз (Matthew Collins) говорит, что очень долго искренне не верил, что восстановить древние белковые последовательности вообще возможно.
В 2000-х годах в этих целях попробовали использовать масс-спектрометрию, которая уже успешно применялась для анализа современных протеинов.
Методику, обозначаемую ZooMS,стали использовать для определения, к каким животным относятся найденные археологами костные фрагменты.
В этом подходе анализируются конкретные формы коллагена,которые у разных видов животных имеют уникальные особенности - получается своего рода хи-мический «отпечаток пальца», по которому можно определить вид живого су-щества.Именно с помощью ZooMS среди тысяч костных обломков,найденных в Денисовой пещере, обнаружили фрагмент человеческой кости,как выяснилось, принадлежавший гибриду неандерталки и денисовца.Сейчас через эту методи- ку прогоняют 40 тыс. неопознанных костей из Азии, в надежде наткнуться на новых гоминин.
Но ZooMS имеет низкую разрешающую способность. После того как кость определена как человеческая, нужно задействовать более точные методы.
В процессе протеомного анализа в образце стараются «поймать» любые бел-ки - а их могут быть тысячи. Затем всё найденное прогоняют по специальным базам данных, в которых хранятся последовательности коллагена и других белков известных организмов. Сходства и различия в составе аминокислот позволяют выяснить родственные связи древнего животного.
Исследователи надеются, что в ближайшем будущем именно такой подход по-зволит уточнить происхождение загадочного Homo floresiensis, останки которого были обнаружены на индонезийском острове Флорес в 2003 году. Чей потомок флоресский хоббит — яванских питекантропов, или же гораздо более древних австралопитеков? Мэтью Коллинз уверен, что в останках хоббита сохранился белок, а значит, есть материал для исследования.
То же самое можно сказать и о другом недавно описанном человечке, Homo luzonensis с острова Лусон на Филиппинах. Исследователи планируют начать пока что с зуба животного из пещеры, где нашли лусонского карлика, чтобы проверить жизнеспособность методики в тропических условиях.
Пока учёные готовятся к протеомному анализу древних людей, исследование останков других животных уже дало интересные результаты. Энрико Каппелли- ни (Enrico Cappellini), специалист по палеопротеомике из Копенгагенского уни-верситета (дат.Københavns Universitet),использовал новый подход для анализа останков ископаемого носорога Stephanorhinus — возрастом 1,8 млн лет!
Аминокислотный состав говорит о тесных связях животного с шерстистым но-сорогом Coelodonta antiquitatis (сейчас в Nature готовится к публикации статья об этом исследовании). Замечательно место, из которого происходят останки Stephanorhinus - Дманиси в Грузии! Вы подумали о том же,о чём я? Если полу- чилось с носорогом,следующий на очереди - «Человек грузинский» из Дманиси.
Интересно, что в этом случае белок добыт не из дентина, а из зубной эмали. Эмаль — самый твёрдый материал в нашем организме, и образует замкнутую систему, в которой белки должны очень хорошо сохраняться. По словам исследователей, почти 2 млн лет — это далеко не предел!
Другие учёные пошли ещё дальше. Сообщают об извлечении следов коллагена из останков верблюда возрастом 3,4 миллиона лет, обнаруженных в Арктике.
А в исследовании 2016 года итальянские специалисты получили белок из яич-ной скорлупы страуса, жившего 3,8 миллиона лет назад. Причём находка сде-лана в Танзании, где среднегодовая температура составляет около 18 °C. Это внушает надежду на успешное исследование костей африканских гоминин.
Конечно, на пути исследователей возникает немало трудностей. Коллаген из тибетской челюсти соотносили с генетическими последовательностями неан-дертальцев и сапиенсов, а с кем сравнивать австралопитеков? У нас нет ДНК такой древности!
Сванте Паабо (швед. Svante Pääbo), самый известный палеогенетик в мире, говорит и о другой серьёзной проблеме - загрязнении образцов посторонней органикой. На то, чтобы научиться отличать древнюю ДНК от современной, у генетиков ушли годы.Но прежде исследователи успели опубликовать немало сенсационных результатов, которые не прошли проверки и оказались в итоге ложными - вроде прочтения ДНК динозавров и древних насекомых, застывших в янтаре.
Другие ограничения более фундаментальны. В ископаемых останках остаётся очень мало белка. Из челюсти тибетского денисовца исследователям удалось получить последовательности 8 видов коллагена, что составляет в сумме чуть более 2000 аминокислот. Только одна из этих аминокислот отличалась от неан- дертальских и сапиентных вариантов,что и позволило отнести находку к дени- совцам. Могло и не повезти! Выбрали бы для анализа белки,содержащиеся не в дентине, а в зубной эмали - и отличий не было бы вовсе. Сравните с ДНК: один древний геном может содержать порядка 3 млн отличий от другого.
Специалисты уверены, что все препятствия удастся преодолеть. Чем древнее животное,тем сильнее состав его белков должен отличаться от современного, так что сравнительный анализ будет более информативным. Важно не созда-вать вокруг новых открытий нездоровый шум, и спокойно оттачивать методику, для чего необходимы согласованные усилия учёных разных специализаций.
Среди таких исследователей Джессика Хенди (Jessica Hendy), археолог из Йоркского университета (University of York),Великобритания,которая использует протеомный анализ для изучения рациона древних людей. В публикации 2018 года Хенди и её коллеги идентифицировали белки, сохранившиеся на поверх-ности 8000-летней керамики из Турции. Удалось установить, какие растения и каких животных употребляли в пищу хозяева древней посуды, и даже выяснить, что они перерабатывали молоко в сыворотку.
Хотя для газетных заголовков привлекательней всего тема эволюции человека, учёные задействуют протеомику для самых разных целей: от исследования древних заболеваний зубов до анализа шкур животных, из которых сделаны средневековые пергаменты.
Keskustelua:
http://keskustelu.skepsis.fi/Message/FlatMessageIndex/89955?page=1#90123
RK
06.09.2002 02:45:55
89955
Steven Pinker vauhdissa jälleen
Mahtaakohan tuo yksi ja sama sanoma jankuttamalla parantua vai huonontua:
http://www.newscientist.com/opinion/opbooks.jsp?id=ns235923
Taas saavat Pinkeriltä kyytiä muutenkin ja paremmin perustein tuhanteen kertaan kumotut Skinnerit, Freudit ja Kaganit, muttei henkäystäkään niistä Pinkerismin-Chomskismin-Wilsonismin vastustajista, joita _ei ole_ kumottu...
RK
Muokannut: , 6/7/2012 7:38:23 PM
RK
09.09.2002 00:00:51
90051
Re: Steven Pinker vauhdissa jälleen
748 kirjoitti 09.09.2002 (90042)...
>RK kirjoitti 06.09.2002 (89955)...
>>Mahtaakohan tuo yksi ja sama sanoma jankuttamalla parantua vai huonontua:
>>http://www.newscientist.com/opinion/opbooks.jsp?id=ns235923
>>Taas saavat Pinkeriltä kyytiä muutenkin ja paremmin perustein tuhanteen kertaan >>kumotut Skinnerit, Freudit ja Kaganit, muttei henkäystäkään niistä Pinkerismin- >Chomskismin-Wilsonismin vastustajista, joita _ei ole_ kumottu...
>>RK
>Pinkerismi...? Ettei vaan unohtuisi värilliset silmälasit päähän kun henkilön nimeen >liitetään -ismi?
Kaikilla on jonkin väriset silmälasit, esimerkiksi Pinkerillä kuten minullakin. Pinkerin lasien poliittinenkin väri tuosta jutusta huomattavasti selkenee, mikä on itse asiassa eräänlaista edistystä...
RK
RK
09.09.2002 00:01:03
90063
Lisää lunta tupaan Pinkerille...
Ja tässä sitä yllämainittua tulee.
http://www.nature.com/cgi-taf/DynaPage.taf?file=/nature/journal/v419/n6902/full/419019a_fs.html
Juuri Pinkerin yhtenä keskeisenä metodina todistaa "synnynnäistä ajattelua" ovat olleet nuo pikkulasten yllättyneet ilmeet ja pitkät katseet "järjenvastaisen", "epäratio-naalisen", "odottamattoman" ilmiön edessä: esimerkiksi verhon taakse menee kaksi Mikki-hiirtä, mutta kun verho poistetaan niitä onkin siellä kolme tmv. Samoin ne ovat olleet keinona "todistaa" eläinten muka rationaalista tai "loogista" ajattelua. Kuitenkin tuollainen "epärationaalisen" tai "järjenvastaisen" havaitseminen edellyttäisi käsittei- tä, joita paljon isommillakaan lapsilla ei suinkaan aina ole, ei opittuina eikä luonnos-taan, vaan yksi ja toinen aivan mahdoton, taatusti epärationaalinen ilmiö menee ns. läpi kuin tyhjää vaan (eivätkä he silti missään tapauksessa ole "tyhmempiä" kuin puolivuotiaat).
Silti sellaisella käyttäytymisellä, joka vaikuttaa yllättyneisyydeltä, saattaa olla teke-mistä oppimisen kanssa: ei esimerkiksi löydy ehdollistunutta aiempaa ketjua, johon havainto liittyisi, ja jollaisista ketjuista sitten myöhemmissä kehitysvaiheissa abstra-hoituisi ´rationaalisen´ (so aiemman kokemuksen kanssa sopusoinnussa olevan) käsite.
...Aivoistamme näyttää pikemminkin puuttuvan joitakin vielä apinoillakin ominaisia piirteitä, jotka ovat olleet esimerkiksi kilen kehittymisen esteenä, kuin että siellä olisi joitakin periaatteellisesti aivan uudenlaisia monimutkaisia synnynnäisiä rakenteita verrattuna "serkkuihimme"...:
http://health.ucsd.edu/news/2002/08_26_Varki.html
http://www.reuters.com/news_article.jhtml?type=sciencenews&StoryID=1377121
Se on ollut aina pääteltävissä ehdollistumisteorian pohjalta, mutta nyt ruvetaan saamaan myös konkreettista tietoa.
https://notes.utk.edu/bio/greenberg.nsf/0/ae1ad66f5a60a9bf85256c30003a0c38?OpenDocument
http://www.nybooks.com/articles/archives/2003/feb/27/darwinian-storytelling/
RK
RK
10.09.2002 00:02:03
90123
Re: Mitä tässä lukikaan?
Virpi Kauko kirjoitti 10.09.2002 (90112)...
>RK kirjoitti 10.09.2002 (90100)...
>>Aivoistamme näyttää pikemminkin puuttuvan joitakin vielä apinoillakin ominaisia >>piirteitä,jotka ovat olleet esimerkiksi kilen kehittymisen esteenä,kuin että siellä olisi >>joitakin periaatteellisesti aivan >>uudenlaisia monimutkaisia synnynnäisiä >>rakenteita verrattuna "serkkuihimme"...:
>Luetun ymmärtämisesi tökkii taas aika >pahasti. Antamissasi linkeissä kerrotaan, >että ihmiseltä on kadonnut tietty geeni, joka useimmilla muilla nisäkkäillä, >ihmisapinat mukaanlukien, on.
>Kyseinen geeni tuottaa tietynlaista sokeria solujen pinnalle. Siis ihmiseltä puuttuu >tuo GEENI ja tuo SOKERI, ei mitään aivojen piirteitä. Tutkijat vain arvelevat että >mutaatiolla SAATTAA olla jotain tekemistä aivojen laajenemisen kanssa koska nuo >asiat ovat tapahtuneet samoihin AIKOIHIN. Eli tämäkään mielenkiintoinen uutinen >ei millään tavalla tue luulojasi.
Tällä uskotaan olevan Ajit Varkin mukaan olevan tekemistä tuon aivojen muovautu-vuuden eli osapuillen paremman oppimiskyvyn kanssa. Tieto on myös 7.9. 2002 Hesarissa. Se on tainnut lakta tasnssimasta "Darwin"-seuran pillin mukaan ainakin yhdeksi päiväksi... Tuo sialiini on itse asiassa vanha juttu, mutta ensimmäistä kertaa näen asiasta suomalaisessa lehdessä.
Mitä tulee aivojen laajenemiseen, niin siitäkin on kyllä uutta tietoa:
http://www.sciencemag.org/cgi/content/full/297/5580/365
Hiirelle saadaan poimuttunut laajempi aivokuori erilaistumattomista soluista tietyllä beetakateiinilla. Tämäkään ei kauheasti todista, että siellä aivokuoressa olisi kovin valmista käyttäytymisen "työkalupakkia" hiirelläkään ainakaan solujen valmiisiin yh-teyksiin koodattuna (muualla aivoissa niitä kyllä hiirellä voi olla vaikka kuinka paljon) ...
RK
Virpi Kauko
11.09.2002 00:02:33
90153
Kromosomien määristä
M-P.Hirvonen kirjoitti 10.09.2002 (90134)...
>muistelen, että esim. simpanssilla on kokonainen kromosomi enemmän kuin >ihmisellä.Mitä se sitten sisältäneekään,ehkäpä 98% samoja geenejä kuin ihmisellä.
>98:n ??
Muistelet oikein, mutta tuon "ylimääräisen" kromosomin sisältämä perimä ei silti kokonaan puutu ihmiseltä.
Tietyt kaksi kromosomia,jotka simpanssilla ovat eri (samoin muillakin ihmisapinoilla), ovat ihmisellä liittyneet yhdeksi pötköksi.
Täällä on pitkä juttu hominidien evoluutiosta, sivun loppupuolella hyvä kaaviokuva kromosomeista:
http://www.micro.utexas.edu/courses/levin/bio304/humanevol/humanevol.html
Täältä löytyy asiaa koskevia harjoitustehtäviä, ilmeisesti tarkoitettu biologian opiskelijoille:
http://www.indiana.edu/~ensiweb/lessons/chr.com2.pdf
ja täällä lisää selityksiä:
http://www.madsci.org/posts/archives/may2001/989331026.Ev.r.html
http://keskustelu.skepsis.fi/Message/FlatMessageIndex/96433?page=2#96548
RK
08.11.2002 01:49:08
96548
Tieteellinen ihmiskuva syvenee nopeaa vauhtia.
Ism kirjoitti 07.11.2002 (96433)...
>Uuden tieteellisen tutkimuksen mukaan ihmisen ja ja simpanssin DNA:n ero on >huomattavasti suurempi kuin aikaisemmin on väitetty. Samoja geenejä onkin uuden >tutkimuksen mukaan alle 95 prosenttia, kun aikaisemmat tutkimukset ovat >päätyneet n. 98,5% %:in samankaltaisuuteen. Evoluutioteorian todisteeet näyttävät >selvästi murentuvan yksi toisensa jälkeen.
http://www.newscientist.com/news/news.jsp?id=ns99992833
Ero varmasti vielä kasvaa, jos otetaan huomioon, että eri pätkät yhteisestäkin DNA:sta kuuluvat aktiiviseen tai hiljaiseen DNA:han eri lajeilla.
Mutta itse evoluutioteorian pätevyyteen tuo ei vaikuta pätkääkään.
Suurimman eron ihmisen ja simpanssin välille aiheuttaa, että solupintojen sokerit poikkeavat melko jyrkästi lajien välillä. Simpanssin sialiini (siaalihappo, sialic acid) on korvautunut ihmisellä suurella joukolla monimuotoisempia sokeriyhdisteitä. Juuri tämän arvelee Kalifornian San Diegon yliopiston Sokeribiologian tutkimuskeskuksen johtaja professori Ajit Varki olevan ihmisen aivokuoren simpanssiin nähden suunnat- tomasti tehokaamman ehdollistumiskapasiteetin taustalla New Scinentistissa 26.1 0. 02 (Sugar Rush: Did sugars Make us smart?). Valitettavasti New Scientistin tämän alan jutut eivät ole nykyään netissä.
Kokeet osoittavat, että sellaisilla eläimillä kuten hiirillä ja apinoilla sialiinitaso on aivoissa paljon matalampi kuin muissa kudoksissa, mutta vain ihmisellä kyseinen aine puuttuu kokonaan,ja se puuttui myös neandetalin ihmiseltä. Varki ja muut val-mistelevat kokeita, joissa hiiren aivoista poistetaan sialiini, ja katsotaan mitä ilmenee (aivan automaattisesti ei heti välttämättä ilmene mitään, mutta myöhemmin voidaan ehkä monimutkaistaa muiden sokerien valkoimaa).
Sokerit toimivat erilaisina "kynsinä", "kytkiminä" ja "tunnistusnappeina" kaikkienkin solujen pinnalla, mutta aivokuorella niillä näyttäisi olevan aivan erityinen rooli juuri ehdollistumisjärjestelmässä. Niillä on nimittäin geenien koodaamiin valkaisaineisiin nähden suunnattomasti suurempi monimuotoisuus ja sitä tietä potentialinen informaatiokapasiteetti. Professori Ajit Varki kirjoittaa mainitussa lehdessä:
"Jos kysytte, mikä on tietyn soltyypin GLYKOMI (analoginen sen genomille ja proteo-mille, kaikkien proteiinityyppien joukolle), niin se on monituhatkertaisesti se genomia monimutkaisempi." (s. 35)
Solussa monimutkaisia sokereita syntetisoi ns. Golgin laite entsyymien avulla, jotka ovat geenien määräämiä. Geenit eivät siis suoraan koodaa sokereita,ja välillisestikin ne voivat periaatteessakaan määrätä eksakstisti vain joitakin tärkeimpiä yksinkertai-sia sokereita. Lopulisen muotonsa monimutkaiset molekyylit saavat ympäristö- ja solunulkoisistakin syistä erityisesti solujen pinnalla.
Sokerit koostuvat noin kymmenestä "alkeispalasta", kuten geenit koostuvat kahdesta (adeniini-tymiini ja sytosiini-guamiini, vai miten se meni, jotka lisäksi voivat olla kah-della tavalla ketjussa). Kaksi noista kymmenestä on jokaiselle tutut glukoosi (rypäle-sokeri) ja fruktoosi (hedelmäsokeri, ja niiden yhdistelmä on sitten tavallinen ruokoso-keri).Nuo palat,kaksoisrenkaat voivat kuitenkin liittyä toisiinsa monin eri tavoin, kuten erilaisissa kulmissa. Kuvavaa on, että kun kuuden askelman DNA-pätkällä on 4^6 = 4096 mahdollista muotoa, niin kuuden alkeissokerirenkaan glykosaminoglykaanilla mahdollisia muotoja on 12 miljardia (12 billions). Sokereiden yhdistämisestä valku-aisaineiden tai rasvojen pinnalle huolehtivat glykosyylitransferaasientsyymit, joiden toimintaa voidaan jossakin määrin hallita. On mm. kehitetty rokote, joka tekee immuuniksi tiettyä MYRKKYÄ (ei siis jotakin tiettyä pöpöä) vastaan.
Eikä niin hyvää ettei jotakin "pahaakin": ruotsalaisetkin heiluvat taas pitkästä aikaa mukana tieteellisessä kärjessä ja Uumajan (ei kuitenkaan, huhhuh, Upsalan..) yliopistossa juui satiin identifioitua sokerireseptori, joka ratkaiseen pääseekö helikobakteeri infektoimaan mahaa vai ei...
Hyvästi "kvanttitietoisuus" ja hyvästi "sosiobiologia/evoluutiopsykologia", ei tule ikävä.
RK
***
Nykyihmisen ja neandertalilaisen sialiinimutaationjälkeiset aivokemikaalit poikkeavat yhden proteiinin TKTL1 osalta
https://scitechdaily.com/key-differences-revealed-between-brains-of-modern-humans-and-neanderthals/
" Key Differences Revealed Between Brains of Modern Humans and Neanderthals
TOPICS: Brain Max Planck Institute Neanderthals Neuroscience Popular
By Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG) September 9, 2022

Scientists uncover a greater neuron production in the frontal lobe during brain development in modern humans than Neanderthals due to the change of a single amino acid in the protein TKTL1.
What makes modern humans unique? It is a question that has long been a driving force for researchers. Therefore, fascinating insights are revealed by comparisons with our closest relatives,the Neanderthals. The increase in brain size and increased neuron production during brain development are considered to be primary factors for the increased cognitive abilities that occurred during human evolution.
However, while both Neanderthals and modern humans develop brains of similar size, very little is known about whether modern human and Neanderthal brains may have differed in terms of their neuron production during development.
Scientists from the Max Planck Institute of Molecular Cell Biology and Genetics (MPI - CBG) in Dresden have discovered that the modern human variant of the protein TKTL1, which differs by only a single amino acid from the Neanderthal variant, in-creases a specific type of brain progenitor cells,called basal radial glia,in the modern human brain.Basal radial glial cells generate the majority of the neurons in the deve- loping neocortex, a part of the brain that is essential for many cognitive abilities.
Because TKTL1 activity is particularly high in the frontal lobe of the fetal human brain, the scientists conclude that this single human-specific amino acid substitution in TKTL1 underlies a greater neuron production in the developing frontal lobe of the neocortex in modern humans than in Neanderthals.
Just a small number of proteins have differences in the sequence of their amino acids – the building blocks of proteins – between modern humans and our extinct relatives, the Neanderthals and Denisovans. It is largely unknown what the biological significance of these differences is for the development of the modern human brain. In fact, modern humans and Neanderthals feature a brain, and notably a neocortex, of similar size, but whether this similar neocortex size implies a similar number of neurons remains unclear.

Microscopy picture of a dividing basal radial glial cell, a progenitor cell type that generates neurons during brain development. Modern human TKTL1, but not Neanderthal TKTL1, increa-ses basal radial glia and neuron abundance. Credit: Pinson et al., Science 2022 / MPI-CBG
The latest study of the research group of Wieland Huttner, one of the founding directors of the Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG) in Dresden, addresses just this question. The research was carried out in collaboration with Svante Pääbo, director at the Max Planck Institute for Evolutionary Anthropology in Leipzig, and Pauline Wimberger of the University Hospital Dresden and their colleagues.
The scientists focus on one of these proteins that presents a single amino acid change in essentially all modern humans compared to Neanderthals, the protein transketolase-like 1 (TKTL1). Specifically, in modern humans, TKTL1 contains an arginine at the sequence position in question, whereas in Neanderthal TKTL1 it is the related amino acid lysine. In the fetal human neocortex, TKTL1 is found in neo-cortical progenitor cells, the cells from which all cortical neurons derive. Notably, the level of TKTL1 is highest in the progenitor cells of the frontal lobe.
New research reveals that a change in a single amino acid in the protein TKTL1 results in greater neuron production in the developing frontal lobe of the neocortex in modern humans compared with Neanderthals.
Anneline Pinson, the lead author of the study and researcher in Wieland Huttner’s group, set out to investigate the significance of this one amino acid change for neocortex development. Anneline and her colleagues introduced either the modern human or the Neanderthal variant of TKTL1 into the neocortex of mouse embryos.
They observed that basal radial glial cells,the type of neocortical progenitors thought to be the driving force for a bigger brain, increased with the modern human variant of TKTL1 but not with the Neanderthal variant. As a consequence, the brains of mouse embryos with the modern human TKTL1 contained more neurons.
More neurons in the frontal lobe of modern humans
After this, the investigators explored the relevance of these effects to human brain development. To this end, they replaced the arginine in modern human TKTL1 with the lysine characteristic of Neanderthal TKTL1, using human brain organoids. These are miniature organ-like structures that can be grown from human stem cells in cell culture dishes in the lab and mimic aspects of early human brain development.
“We found that with the Neanderthal-type of amino acid in TKTL1, fewer basal radial glial cells were produced than with the modern human-type and, as a consequence, also fewer neurons,” says Anneline Pinson. “This shows us that even though we do not know how many neurons the Neanderthal brain had,we can assume that modern humans have more neurons in the frontal lobe of the brain, where TKTL1 activity is highest, than Neanderthals.”
The researchers also discovered that modern human TKTL1 acts through changes in metabolism. Specifically, stimulation of the pentose phosphate pathway followed by increased fatty acid synthesis. In this way, modern human TKTL1 is thought to increase the synthesis of certain membrane lipids needed to generate the long process of basal radial glial cells that stimulates their proliferation and, therefore, increases neuron production.
“This study implies that the production of neurons in the neocortex during fetal deve-lopment is greater in modern humans than it was in Neanderthals, in particular in the frontal lobe,” summarizes Wieland Huttner, who supervised the study. “It is tempting to speculate that this promoted modern human cognitive abilities associated with the frontal lobe.”
Reference: “Human TKTL1 implies greater neurogenesis in frontal neocortex of modern humans than Neanderthals” by Anneline Pinson, Lei Xing, Takashi Namba, Nereo Kalebic, Jula Peters, Christina Eugster Oegema, Sofia Traikov, Katrin Reppe, Stephan Riesenberg, Tomislav Maricic, Razvan Derihaci, Pauline Wimberger, Svante Pääbo and Wieland B. Huttner, 9 Sepetember 2022, Science.
DOI: 10.1126/science.abl6422 "
TKTL1 and hominin cortical neurogenesis.

The single lysine-to-arginine substitution in modern human TKTL1 leads to greater bRG num-bers than in Neanderthals. These bRG in turn generate more neocortical neurons in modern hu-mans. Because TKTL1 expression in fetal human neocortex is particularly high in the developing frontal lobe, these findings imply that the frontal lobe of modern humans contains more neurons than that of Neanderthals.
" Neanderthal brain development
Neanderthal brains were similar in size to those of modern humans but differed in shape. What we cannot tell from fossils is how Neanderthal brains might have dif-fered in function or organization of brain layers such as the neocortex. Pinson et al. have now analyzed the effect of a single amino acid change in the transketolase-like 1 (TKTL1) protein on production of basal radial glia, the workhorses that generate much of the neocortex (see the Perspective by Malgrange and Nguyen). Modern humans differ from apes and Neanderthals by this single amino acid change. When placed in organoids or overexpressed in nonhuman brains, the human variant of TKTL1 drove more generation of neuroprogenitors than did the archaic variant. The authors suggest that the modern human has more neocortex to work with than the ancient Neanderthal did. — PJH
Structured Abstract
INTRODUCTION
The evolutionary expansion of the neocortex and the concomitant increase in neuron production are considered to be a basis for the increase in cognitive abilities that oc-curred during human evolution. Endocast analyses reveal that the endocranial vo-lume of modern humans and Neanderthals was similar, suggesting similar brain and neocortex size. But whether similar neocortex size implies similar neocortical neuron production remains unclear.
RATIONALE
Transketolase-like 1 (TKTL1) is a gene from the transketolase family that in fetal human neocortex is preferentially expressed in the two classes of neuroprogenitors, the apical progenitors in the ventricular zone and the basal progenitors in the sub-ventricular zone. The latter class of neuroprogenitors comprises two major types, the basal intermediate progenitors (bIPs) and the basal radial glia (bRG,also called outer radial glia). bRG exhibit cellular processes that promote their ability to self-amplify, and are the neuroprogenitor type considered to be a driver of the increase in cortical neuron production, which is a hallmark of the evolution of the human neocortex.
Reflecting their cell polarity, bRG undergo repeated asymmetric divisions that self-renew the bRG and generate one neuron each. Thereby, bRG generate more neurons over time than the other type of neuron-generating basal neuroprogenitors, the process-lacking bIPs whose neurogenic divisions are symmetric self-consuming.
TKTL1 is one of the few proteins with a single amino acid substitution found in essentially all present-day humans but absent from extinct archaic humans, the Neanderthals and Denisovans, and other primates. This human-specific amino acid substitution in TKTL1 is a lysine in apes and archaic humans but an arginine in mo- dern humans. We therefore investigated (i) whether TKTL1 has a role in neocortex development and affects neuroprogenitor numbers and (ii) whether both archaic TKTL1 (aTKTL1) and modern human TKTL1 (hTKTL1) exert similar effects on neuroprogenitors during neocortex development.
RESULTS
When expressed in mouse embryo neocortex, which lacks TKTL1 expression, hTKTL1 increased the abundance of bRG without affecting that of bIPs and that of apical progenitors. The effect of TKTL1 on bRG abundance was limited to hTKTL1; aTKTL1, which differs only by one amino acid, was unable to increase bRG abundance. The greater bRG abundance upon hTKTL1 expression resulted in an increase in cortical neuron production over time, specifically of the late-born upper-layer neurons rather than of the early-born deep-layer neurons. In the folded (gyren-cephalic) developing ferret neocortex, hTKTL1 expression increased not only bRG abundance but also the proportion of bRG with multiple processes, a hallmark of bRG that can self-amplify. As a consequence of this effect, gyrus size increased.
In fetal human neocortex, hTKTL1 was essential to maintain the full number of bRG, as CRISPR-Cas9–mediated hTKTL1 knockout in fetal human neocortical tissue re-duced this number. To further demonstrate the relevance of this effect, we converted hTKTL1 to the Neanderthal variant aTKTL1 in human embryonic stem cells and ge-nerated minibrain structures called cerebral organoids. The aTKTL1-expressing or-ganoids contained fewer bRG and neurons, hence the human-specific lysine-to-argi- nine substitution in hTKTL1 is essential for maintaining the full number of bRG and neurons in this human brain model. In fetal human neocortex, hTKTL1 expression in neuroprogenitors increased during the course of neurogenesis and was particularly high in the developing frontal lobe as compared to the developing occipital lobe.
As to its mechanism of action, hTKTL1 increased bRG abundance via two metabolic pathways, the pentose phosphate pathway (PPP) followed by fatty acid synthesis. Inhibition of the PPP or of fatty acid synthesis, using a variety of specific inhibitors, completely suppressed the hTKTL1-induced increase in bRG abundance in embryo-nic mouse neocortex and reduced bRG numbers in fetal human neocortical tissue. This metabolic action of hTKTL1, but not aTKTL1, in bRG resulted in an increase in the concentration of acetyl–coenzyme A, the critical metabolite for fatty acid synthe-sis. Our data suggest that hTKTL1, but not aTKTL1, promotes the synthesis of mem-brane lipids containing a certain type of fatty acid that are required for the outgrowth of bRG processes and hence for the increase in bRG abundance.
CONCLUSION
In light of our finding that TKTL1 expression in fetal human neocortex is particularly high in the developing frontal lobe, our study implies that because of the single amino acid–based activity of hTKTL1, neocortical neurogenesis in modern humans was and is greater than it was in Neanderthals, in particular in the frontal lobe. "
https://www.mpi-cbg.de/research/researchgroups/currentgroups/wieland-huttner/research-focus

Wieland B. Huttner on "Which Evolutionary Changes in the Genome Led to the Development of the Large-Sized Human Brain?"
" Wieland Huttner Group
MPI-CBG > Research > Research Groups > Current Groups > Wieland Huttner > Research Focus
Neural stem and progenitor cells and neocortex expansion in development and evolution
Our goal is to elucidate the molecular and cellular mechanisms underlying the evolu-tionary expansion of the neocortex, specifically the increase in the number of cortical neurons generated during embryonic/fetal development. Ultimately, we aim at identi-fying the genomic changes that are responsible for the increase in neuron number in the human neocortex as compared to other primates.
Towards this goal, we take the following approaches.
1. We characterize neural stem and progenitor cells (SPCs) that generate neurons in the developing neocortex of a variety of species (mouse, ferret, marmoset, human), and determine their lineage.
https://en.wikipedia.org/wiki/Neural_stem_cell
https://en.wikipedia.org/wiki/Progenitor_cell
Using this knowledge,we study the differences across mammals developing a lissen- cephalic versus gyrencephalic neocortex, in order to obtain insight into neocortex evolution.
2. We investigate specific cell biological features of the various cortical SPCs, inclu-ding their cell polarity and organization and dynamics of intracellular organelles, in order to obtain clues as to their differential proliferative potential.
3. We study the cell cycle and cell division of cortical SPCs.
4. We explore the role of genes, mutations in which cause microcephaly,and develop novel technology to functionally search for, and test, candidate genes for neocortex expansion.
5.We analyze the transcriptomes of cortical germinal zones and specific SPC subpo- pulations,in order to identify,and then functionally test,candidate genes for neocortex expansion.
Documentary of the MDR "Accidentally intelligent? The small step to a big brain” from June 2020, featuring the Wieland B. Huttner and Michael Heide and their research. ENGLISH version
Documentary of the MDR "Accidentally intelligent? The small step to a big brain” from June 2020, featuring the Wieland B. Huttner and Michael Heide and their research. GERMAN version "
***
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC21712/
A mutation in human CMP-sialic acid hydroxylase occurred after the Homo-Pan divergence
Abstract
Sialic acids are important cell-surface molecules of animals in the deuterostome lineage. Although humans do not express easily detectable amounts of N-glycolyl-neuraminic acid (Neu5Gc, a hydroxylated form of the common sialic acid N-acetyl-neuraminic acid, Neu5Ac), it is a major component in great ape tissues, except in the brain.This difference correlates with lack of the hydroxylase activity that converts CMP-Neu5Ac to CMP-Neu5Gc. Here we report cloning of human and chimpanzee hydroxylase cDNAs. Although this chimpanzee cDNA is similar to the murine homo-logue, the human cDNA contains a 92-bp deletion resulting in a frameshift mutation.
The isolated human gene also shows evidence for this deletion. Genomic PCR ana-lysis indicates that this deletion does not occur in any of the African great apes. The gene is localized to 6p22–p23 in both humans and great apes, which does not corre-spond to known chromosomal rearrangements that occurred during hominoid evolu-tion. Thus,the lineage leading to modern humans suffered a mutation sometime after the common ancestor with the chimpanzee and bonobo, potentially affecting recog-nition by a variety of endogenous and exogenous sialic acid-binding lectins.Also, the expression of Neu5Gc previously reported in human fetuses and tumors as well as the traces detected in some normal adult humans must be mediated by an alternate pathway.
Keywords: Pongidae/evolution/neuraminic acids/PCR/hominoids
Humans are evolutionarily related to the African great apes, Pan troglodytes (the chimpanzee), Pan paniscus (the bonobo or pygmy chimpanzee) and Gorilla gorilla (the gorilla) (1–6). Current data indicate that the last common ancestor of humans with the great apes was very likely shared with the chimpanzee and bonobo (7–16). Given the ≈99% identity at the DNA and amino acid-sequence level, it is proposed that only a few changes in gene expression or function are responsible for the mor-phological and functional differences between humans and these closely related pri-mates (4). Such differences in gene regulation and function could also explain varia-tions in susceptibility and biological response to diseases such as cancer, hepatitis, AIDS, malaria, and intestinal infections (17–22).
The sialic acids are a family of acidic sugars typically found at the outer end of the cell surface and secreted glycoconjugates of all vertebrates (23–26). The two most common forms of sialic acid found in mammalian cells are N-acetylneuraminic acid (Neu5Ac) and its hydroxylated derivative, N-glycolylneuraminic acid (Neu5Gc). The conversion from Neu5Ac to Neu5Gc can positively or negatively affect interactions involving several of the known endogenous and exogenous receptors for sialic acids such as CD22,myelin-associated glycoprotein,sialoadhesin,and the influenza A virus hemagglutinin (27–33). Interestingly, even in animals with large amounts of Neu5Gc in other tissues,its level in the brain is always extremely low (34–36). We recently re- ported a major biochemical difference between the great apes and humans, with the latter showing a loss of activity of the enzyme CMP-N-acetylneuraminic acid hydro-xylase (37).This enzyme converts the nucleotide sugar donor CMP-Neu5Ac to CMP-Neu5Gc in other animals (38–46). Loss of hydroxylase activity provides a potential explanation for the finding that although the body fluids and tissues of all of the great apes (chimpanzee, bonobo, gorilla, and orangutan) express high levels of this sialic acid, corresponding samples from humans contain little or no detectable Neu5Gc (37).
However,the regulation of this hydroxylase activity is very complex and involves mul-tiple interacting factors (38-45). Moreover,expression of Neu5Gc has been reported in human fetal tissues (47) and in certain human cancers (47–49), and our previous study (37) noted that some human tissues had small quantities of an HPLC peak corresponding to Neu5Gc. Thus, it has been suggested that the activity of this hydroxylase may be suppressed in adult humans, rather than eliminated altogether.
To explore this issue, we have cloned the human and chimpanzee cDNAs encoding the CMP-Neu5Ac hydroxylase. We find that the human lineage has suffered a gene-tic mutation in the coding region of this cDNA. While this manuscript was in prepara-tion, another group reported the same deletion in a human hydroxylase cDNA (50). However, that paper reported an incomplete cDNA sequence, which did not include the initial 5′ translation start site. It also did not address the uniqueness of this mutation for humans and its implications for hominoid evolution.
MATERIALS AND METHODS
Identification and Characterization of Expressed Sequence Tag (EST) Clones.
The previously reported mouse CMP-Neu5Ac hydroxylase sequence (45) was used to carry out blast searches of the human EST database at the National Center for Biotechnology Information website (http://www.ncbi.nlm.nih.gov/dbEST/index.html). Several clones with >80% homology to the murine hydroxylase sequence were found. These were obtained from the company Research Genetics, and subjected to complete sequencing of both strands by the dideoxy chain-termination method (performed by the MacConnell Research Corporation, or in the laboratory of S. Hedrick, using an Applied Biosystems Prism Sequencer).
5′ RACE (Rapid Amplification of cDNA Ends).
RACE was performed by using primers based on the 3′ regions of the EST clones. Epstein–Barr virus (EBV)-transformed lymphocytes from chimpanzees and humans were cultured in RPMI medium 1640 supplemented with 10% fetal calf serum and 1% l-glutamine. Messenger RNA was isolated from ≈30 million cells by using a Qiagen Oligotex Direct mRNA isolation kit (Chatsworth,CA). Cells were lysed, homo- genized using Qiagen’s QIAshredder, and incubated with Oligotex beads to allow mRNA to hybridize with (dT)30 oligonucleotides covalently linked to the Oligotex beads. The mRNA-Oligotex complex was then washed twice, and mRNA was eluted with a low-salt buffer.GIBCO/BRL’s 5′ RACE system was then used to perform cDNA amplification. Briefly, 2.5 μg of mRNA was reverse-transcribed with a gene-specific primer RACE1 (CTG TGT TTC CAA CAT AAA TGG G) using Superscript II reverse transcriptase. The mRNA was then degraded with RNase, and the cDNA was puri-fied by using GlassMAX spin cartridges. After dC-tailing of the cDNA with terminal deoxynucleotidyltransferase (TdT) and dCTP, the cDNA was PCR-amplified by using the Abridged Anchor Primer (GIBCO/BRL) and a second gene-specific primer, RACE2 (ACA GCC TCT CCA GCC AAT CAG ATG), to obtain a nested product. The cDNA was then further nested by reamplifying using PCR with the Abridged Univer-sal Amplification Primer (GIBCO/BRL) and the primer RACE3 (AAC CAC CAT CCA CGG GCA AAA GCA), and finally TA cloned by using Invitrogen’s Original TA Cloning Kit. Sequencing of the products was carried out as above.
Cloning of Full-Length cDNA for Chimpanzee and Human Hydroxylase by Reverse Transcription (RT)–PCR.
Two primers, NOTC51 (AAG CAG AGC GGC CGC CAG ACG ATG GGC AGC ATC G) and 31BAM (TGT CTT GGA TCC TTT TCT CTT CCT GTT TCC TC), were desig-ned using 5′ RACE and the 3′ EST sequences common to human and chimpanzee. One microgram of mRNA was reverse-transcribed using 50 pmol of random hexa-mer, 200 units of Superscript II, and 20 units of RNase inhibitor in a 40-μl reaction with First Strand Buffer, 200 μM dNTP, and 10 mM DTT. A 2-μl portion of the reverse transcription reaction was then PCR-amplified with 20 pmol of NOTC51 primer, 20 pmol of 31BAM primer, and the Expand Long Template PCR System from Boehrin-ger Mannheim with 1.75 mM MgCl2. The PCR products were then gel-purified by using the QIAEX Gel Extraction system and cloned by using Invitrogen’s TA Cloning Kit. Sequencing of three independent clones of each PCR product was done as de-scribed above. Final sequences are based on overlapping sequencing runs covering both strands.
Genomic PCR Analysis.
EBV-transformed lymphocytes from human, chimpanzee, bonobo, and gorilla were cultured as described above. Genomic DNA was prepared according to standard procedures (51), and PCR was carried out using primers R5 and R3, corresponding to the 5′ and 3′ ends of the 92-bp region, respectively. Twenty picomoles of R5 (GTC TGT CAG ATG CAC AAA CGA C) and 20 pmol of R3 (CCA GCT CAT CTT GAC AGA AGC) were used to amplify 1 μg of genomic DNA from each species in a 50-μl reaction with 200 μM dNTPs, 2.5 mM MgCl2, 20 mM Tris⋅Cl (pH 8.4), and 50 mM KCl. PCR conditions were as follows: 95°C for 5 min followed by 30 cycles of 94°C, 60 sec; 58°C, 60 sec; 72°C 45 sec, ending with 72°C for 10 min. Ten microliters of the PCR product was then electrophoresed on a 1.5% agarose gel and stained with ethidium bromide.
PCR Amplification of Human 3′ Untranslated Region (UTR) Sequences.
Primers 244MF and 244MR were derived from 3′ UTR sequence of EST244303 (244MF, CAC AGA GGA AAC AGG AAG AG; 244MR, TTG TTA TGC ATG TGA GCG G). PCR was carried out using 20 pmol of each primer and 200 ng of human genomic DNA in a total volume of 20 μl containing 200 μM dNTPs and 1.2 units of Taq polymerase in a buffer of 1.5 mM MgCl2, 15 mM (NH4)2SO4, 60 mM Tris⋅Cl (pH 8.5), and 5% dimethyl sulfoxide. Using a Perkin–Elmer GeneAmp PCR System 9600, the following conditions were used: denaturation at 95°C for 120 sec followed by 32 amplification cycles of 94°C for 30 sec; 58°C for 30 sec; 72°C for 60 sec; and extension at 72°C for 7 min. Electrophoresis of the PCR products through a 2% agarose gel followed by ethidium bromide staining revealed the expected 359-bp product (data not shown).
Isolation of Bacterial Artificial Chromosome (BAC) Clones.
Human genomic clones were identified by screening pools of the human BAC library release III from Research Genetics (Huntsville, AL) by PCR of 3′ UTR sequences. Primers and conditions were as described above, except 100 ng of DNA was used as template. DNA from the positive human genomic BAC clone 443N17 was purified by using a QIAGEN-tip 100 and a modified Plasmid Midi Kit midiprep protocol. Briefly, a 100-ml culture was treated as two 50-ml aliquots until addition to the column. Solutions P1, P2, and P3 were used at 2.5× volumes (compared with the standard protocol) per 50 ml of culture. The column elution buffer QF was heated to 65°C and added to the tip-100 in 1-ml aliquots.
Southern Blot Analysis of BAC Clones.
BAC DNA was prepared, restriction-digested, and analyzed using Southern hybridi-zation performed according to standard procedures (51).BAC DNA (2 μg) was diges- ted overnight at 37°C with BamHI or EcoRI and electrophoresed on a 0.8% agarose gel, which was then shaken for 1 hr in denaturation buffer (0.5 M NaOH/1.5 M NaCl) and neutralized 1 hr in neutralization buffer (1 M Tris⋅Cl, pH 7.5/1.5 M NaCl). The DNA was then transferred for 1 hr 45 min in 20× standard saline citrate (SSC; (1× SSC = 0.15 M sodium chloride/0.015 M sodium citrate, pH 7) onto a nitrocellulose membrane by using a PosiBlot 30–30 pressure blotter (Stratagene) and crosslinked to the membrane by UV irradiation. After rinsing in 5× SSC, the membrane was pre-hybridized overnight at 42°C in prehybridization solution [6× SSC/20 mM NaH2PO4 / 0.4% SDS/5× Denhardt’s solution (1× = 0.02% polyvinylpyrrolidone / 0.02% Ficoll / 0.02% BSA), and 100 μg/ml salmon sperm DNA], and then probed for 72 hr at 42°C with hybridization solution (4× SSC/25 mM NaH2PO4/10% dextran sulfate/4× Den-hardt’s solution/100 μg/ml salmon sperm DNA) containing probes corresponding to the 92-bp region deleted in the human cDNA (PR, based on the chimpanzee se-quence), the sequence immediately 5′ of this region in the cDNA (P5), and the se-quence immediately 3′ of this region (P3). After 3 washes with 2× SSC/0.1%SDS for 1 hr at 42°C, the membrane was exposed onto a PhosphoImager screen (Molecular Dynamics) for 72 hr and then analyzed. Probes were generated by primer extension with α-32P-labeled dNTPs from NEN. Probe PR was generated with primers PR/5 (GCA CAA CTG CAA ATT AGA TGT GAG CAC CAT GAA GTA TAT CAA CC) and PR/3 (CAG AAG CTT TCC GGA GGG TTG ATA TAC TTC ATG GTG);P5 was gene- rated with primers P5/5 (AAG CAC CAA GGA GGC CTG TTC ATA AAA GAT) and P5/3 (CCT CCG GCT AAA TCC TCG ATA TCT TTT ATG); and P3 was generated with primers P3/5 (TTG TTG AAA TGG ATG AAA ACA ACG GAC TTT), P3/3 (GAT TCA GTT CTA AAA GCA AAA GTC CGT TGT).
Human/Rodent Somatic-Cell Hybrid Mapping.
Using 200 ng of DNA from each sample from the NIGMS cell repository human / ro-dent somatic-cell hybrid mapping panel 2 (Coriell Cell Repositories), PCR amplifica-tion and analysis were carried out as above. A radiation-hybrid panel derived from chromosome 6p was screened by using PCR. Primers were designed from the 3′ UTR of the human hydroxylase gene (CHR6.1-F CGA ATC CAA TCA CAG AGG AAA CAG G;CHR6.1-R-GCA TTA TTG TTA TGC ATG TGA GCG G; CHR6.2-F CAA TGG TGG AAT TTG TCT CC; CHR6.2-R CTC ACC ATA GAC ATT CTG ACC TAC; CHR6.3-F CTG AGA TCC TGT TGT GCC TAT CAC C; and CHR6.3-R CCT TGG TGC TTG CAC ATA TTC TTG CAT GCC).
Primer pairs were used as follows: CHR6.1-F/CHR6.1-R; CHR6.2-F/CHR6.2-R; CHR6.3-F/CHR6.3-R; and CHR6.1-F/CHR6.2-R. Primer pair CHR6.3-F/CHR6.3-R produced a PCR product in hamster the same size as the human product. All other primer combinations were informative. PCR was carried out in 25 μl reactions using 100 ng of DNA as template. Reactions were performed in 1× Perkin–Elmer PCR buf-fer/0.1 mM dNTPs (1μM each primer)/5% dimethyl sulfoxide with 1.25 units of Taq polymerase. Amplifications were 30 sec each at 94°C, 57°C,and 72°C, for 30 cycles.
Metaphase Chromosome Preparation.
A transformed normal male lymphoblast cell line was cultured in RPMI medium 1640 supplemented with 10% fetal bovine serum. Cell cultures were treated with Colcemid and harvested by using standard protocols. Fixed-cell suspensions were dropped onto glass slides and were then aged for at least 48 hr at −20°C before use.
Fluorescence in Situ Hybridization (FISH) Analysis.
One microgram of BAC DNA was labeled by nick-translation with digoxigenin 11-dUTP in the presence of reduced dTTP (Boehringer Mannheim), ethanol-precipita-ted, and resuspended in 10 μl of a 65% (vol/vol) formamide hybridization solution. For FISH, 250 ng of labeled DNA was combined with 1 μg of human COT-1 DNA (GIBCO/BRL) to block repetitive sequences. The probe mixture was denatured at 70°C for 10 min and renatured at 37°C for approximately 30 min. FISH was perfor-med according to standard procedures.Slides were dehydrated in ethanol and dena- tured in 70% formamide/2× SSC at 70°C for exactly 2 min. Slides were dehydrated again and hybridized with the BAC 443N17 probe overnight at 37°C in a humidified chamber.The slides were washed in 50% formamide/2×SSC and in 2× SSC at 43°C, and BAC hybridization signals were detected with anti-digoxigenin-rhodamine (Boehringer Mannheim) at 37°C. Chromosomes were counterstained with 4′,6-dia-midino-2′-phenylindole dihydrochloride (DAPI, Boehringer Mannheim) in Vectashield mounting medium (Vector Laboratories), and viewed under a Zeiss Axioskop fluorescence microscope equipped with a triple band pass filter.
RESULTS
Human EST Databases Contain Sequences Homologous to the 3′ Region of the Mouse CMP-Neu5Ac Hydroxylase.
Current human EST databases were found to contain several sequences with 75 – 85% homology to the previously reported mouse CMP-Neu5Ac hydroxylase cDNA. One of these ESTs (clone number 257329) encoded an ORF with as much as 90% identity (with no gaps) to the carboxy-terminal half of the mouse hydroxylase cDNA (see Fig. Fig.1). The1). The only major difference noted is a carboxyl-terminal exten-sion of the ORF that is not present in the mouse cDNA. Two other ESTs (clones 244303 and 701123) contained generally similar sequences, but included some insertions and deletions that prematurely terminate the ORF (data not shown). We reasoned that all of these cDNAs represent alternatively spliced messages derived from the single gene (50) encoding the corresponding human hydroxylase.
Comparison of nucleotide and derived amino acid sequences of CMP-Neu5Ac hyd-roxylase cDNAs from mouse,chimpanzee, and human. (A) Comparison of nucleotide sequences. Sequencing of multiple full-length cDNA clones was carried out as described. The ClustalW program (http://www2.ebi.ac.uk/clustalw/) was used to align the sequences. The 92-bp gap in the human cDNA and differences at the 3′ end are indicated with dotted lines. The start and stop codons of the primary ORF are under-lined. (B) Comparison of the ORFs of the cDNAs. The putative Reiske iron-sulfur binding region of the mouse and chimpanzee hydroxylase are underlined.
Chimpanzee and Human CMP-Neu5Ac Hydroxylase cDNA Homologues Expressed in Lymphoblastoid Cells Are Different in Both the 5′ Region and the 3′ Tail.
We had earlier found that although human B lymphoblastoid cells do not express Neu5Gc, cells from both the bonobo and the chimpanzee do express substantial amounts of this sialic acid. We therefore used the human EST database sequences to obtain complete cDNAs for the chimpanzee and human hydroxylase. We first car-ried out 5′ RACE using mRNA from lymphoblastoid cells and the sequence informa-tion from the 5′ region of the EST clones. The major 5′ RACE product in the human was ≈500 bp in size, whereas the corresponding fragment from the chimpanzee was slightly larger (data not shown). Sequencing indicated that the difference is the result of a 92-bp deletion within the 5′ region of the human cDNA (Fig.(Fig.11A). The 5′ and 3′ sequences were then used to obtain the complete human and chimpanzee hydro-xylase cDNAs by RT-PCR, using a high-fidelity polymerase (Pwo). As shown in Fig. Fig.1,1, the complete chimpanzee cDNA sequence is ≈85% identical to that of the mouse. It includes an ORF that is ≈90% identical to the mouse, with the only major difference being at the 3′ end, where the chimpanzee has a 13-aa extension. In con-trast to the mouse and chimpanzee cDNAs, a 92-bp deletion in the human cDNA results in a frameshift, which would cause a premature truncation of the coding sequence (Fig. (Fig.11B). The deletion includes sequences coding for the putative Reiske iron-sulfur binding region of the murine hydroxylase protein (46), and the re-sulting truncated molecule is therefore highly unlikely to have any hydroxylase activi- ty.This deletion in the cDNA is identical to the one recently reported by Irie et al.(50). However, these authors did not isolate the complete 5′ region of the human cDNA that includes the primary initiator methionine; therefore, they came to a different conclusion regarding the nature of the truncated protein produced (see Discussion).
The cDNA Change in Humans Is Caused by a Genomic Alteration That Is Not Found in Any of the African Great Apes.
Genomic PCR analysis was carried out using primers corresponding to the 5′ and 3′ sequences of the 92-bp region of the chimpanzee cDNA. As shown in Fig. Fig.2,2, we obtained a corresponding 92-bp PCR product from total genomic DNA of all of the African great apes, but not from the human. This suggests that this region of the human genome is either deleted or markedly altered in its sequence. Sequences from the 3′ UTR of human ESTs were therefore used to screen a human genomic BAC array library. PCR screening of BAC pools resulted in the identification of two BAC clones, 443N17 and 541O17. Upon colony purification, DNA of these BAC clones consistently produced an expected 359-bp PCR product. PCR and Southern blot analysis of clone 443N17 with primers and probes representing the 5′ and 3′ ends of the human cDNA indicate that this BAC clone includes the complete coding region for the hydroxylase (data not shown). Further Southern blot analysis with probes corresponding to the sequences immediately 5′ and 3′ of the 92-bp human cDNA deletion indicates that they are present in the BAC clone (Fig. (Fig.3).3).
However, the BAC failed to hybridize with a chimpanzee cDNA probe corresponding to the 92-bp human cDNA deletion (Fig.(Fig.3).3).PCR analysis similar to that shown in Fig. Fig.22 was performed by using the BAC clone as template. Again, no 92-bp product was found, and the higher molecular weight bands seen in the human lane in Fig. Fig.22 were either absent or much less prominent, suggesting that they are nonspecific (data not shown). Further PCR studies (not shown) indicated that the intronic regions on either side of the putative 92 bp-deletion are very large (>10kb) and will require extensive subcloning and sequencing to define precisely.
PCR analysis of genomic DNA from humans and African great apes. Genomic DNA from human (Hu), chimpanzee (Ch), bonobo (Bo), or gorilla (Go) was subjected to PCR analysis with primers corresponding to the 5′ and 3′ ends of the 92-bp sequence missing from the human cDNA. S, molecular weight standards; +, positive control, using chimpanzee cDNA as template; −, negative control, without template. Some higher molecular weight background bands seen with great ape DNA were more prominently amplified in the human sample.
Southern blot analysis of a BAC clone encompassing the human hydroxylase gene. BAC 443N17 DNA (10 μg) was digested with BamHI (HI) or EcoRI (RI) and subjec-ted to Southern blot analysis using probes corresponding to the missing region in the human cDNA (PR,based on chimpanzee sequence),and the sequences immediately 5′ (P5) and 3′ (P3) of this region. All three probes were equally sensitive when used to probe Southern blots of the chimpanzee hydroxylase cDNA (data not shown).
The 92-bp Deletion Is Found in Diverse Human Populations.
Our previous biochemical evidence for Neu5Gc deficiency in humans included a survey of more than 70 individuals from many backgrounds (37). To ensure that the hydroxylase mutation is not simply a human polymorphism, we carried out a similar PCR analyses on DNA samples from 18 “Caucasians,” 4 African Americans, 4 !Kung bushmen, 4 Khwe pygmies, and 6 Japanese. All of these samples failed to give the 92-bp PCR product, whereas control samples from chimpanzees and bonobos showed the product (data not shown).
Chromosomal Localization of the Human Hydroxylase Gene and Comparison with the Great Ape Genes.
A variety of techniques were used to localize the human hydroxylase gene. PCR amplification of human genomic DNA using primers 244MF and 244MR resulted in a PCR product with the expected 359-bp fragment size from the 3′ UTR of EST244303. Amplification of a human/rodent somatic-cell hybrid mapping panel showed the expected product amplified from human genomic DNA and no signal with either mouse or hamster DNA (Fig. (Fig.44A). Only sample 6 also gave a product of 359 bp, indicating that the locus maps to human chromosome 6.
Chromosomal localization of the human CMP-sialic acid hydroxylase gene. (A) Human/rodent somatic-cell hybrid mapping. Ethidium bromide-stained agarose gel analyzing the products of PCR amplification of samples of the NIGMS human/rodent somatic-cell hybrid mapping panel 2. Lanes labeled 1–22, X, and Y reflect DNA from a chromosome 6 hybrid clone containing the designated human chromosomes. Hamster (Ha), mouse (Mo), and human (Hu) are total genomic DNA from those species used to create the hybrid panel. A negative control (B) is shown as well as 100-bp marker lanes with the 300- and 400-bp bands indicated. (B) FISH mapping. Metaphase FISH localization of genomic BAC clone 443N17 containing the CMP-sialic acid hydroxylase gene to the distal short arm of human chromosome 6. Detec-tion of digoxigenin-labeled probes was achieved with anti-digoxigenin antibodies conjugated to rhodamine (giving the red signal), followed by 4′,6-diamidino-2′-phe-nylindole dihydrochloride (DAPI) counterstaining of the chromosomes and viewing under an epifluorescence microscope with a triple band pass filter.
FISH experiments against metaphase chromosomes from a normal human male showed that BAC clone 443N17 maps to the distal short arm of chromosome 6 (Fig. (Fig.44B). PCR primers designed to complement the 3′ UTR of the gene were then used to screen a radiation hybrid panel specific to human 6p (52). This allowed the map position of hydroxylase to be further narrowed to 6p22-p23, to an interval inclu-ding the marker D6S89 and SCA1 (data not shown).FISH studies using chimpanzee, bonobo, and gorilla metaphase chromosomes showed that the map position is con-served with the human (data not shown). Thus, the human hydroxylase gene is not located at the site of any of the gross chromosomal rearrangements known to have occurred in hominoid evolution (53).
DISCUSSION
Our data indicate that the human CMP-Neu5Ac hydroxylase gene underwent a ma-jor mutation since the time of the last common ancestor with the bonobo and chim-panzee. The loss of a 92-bp segment in the coding region results in a frameshift mu-tation that can account for the lack of hydroxylase activity and deficiency of Neu5Gc in humans. We have also isolated the complete human hydroxylase gene, and have shown by Southern blot analysis that it does not contain the 92-bp region. We have not fully characterized the gene because of its very large size. In parallel studies, Irie et al. (50) have reported an analysis of the same region of the human gene, and conclude that a section corresponding to exon 6 of the mouse hydroxylase gene is deleted in humans.
Sialic acids are common structural components of the glycoconjugates of all animals of the deuterostome lineage (23–26) and are known to mediate cell–cell interactions by specific sialic acid-binding lectins (27–31) such as the selectins and the siglecs. A variety of structural modifications of Neu5Ac such as Neu5Gc (23–25) can modulate recognition in endogenous interactions as well as in exogenous microbial recognition (23–26). The expression of Neu5Gc is widespread in animals from sea urchins to mammals, and also shows tissue-specific and developmentally regulated expression (23, 24, 54–56). A change from Neu5Ac to Neu5Gc is likely to affect several of the known endogenous and exogenous receptors for sialic acids (27–29). For example, the presence or absence of Neu5Gc would affect the interactions of microbial patho-gens such as influenza A and B viruses (57–59) and Escherichia coli K99 (60 – 63).
There are many other microbes that utilize sialic acids as specific binding sites on mammalian cells, including major pathogens such as Helicobacter pylori and Plas-modium falciparum (22, 30, 64, 65). In most such instances, the consequences of having Neu5Gc rather than Neu5Ac have not yet been pursued.Because sialic acids are also ligands for a variety of endogenous vertebrate lectins (27–33), the loss of hydroxylation could have complex effects on the growth, development, and function of multiple systems. For example, CD22 on B lymphocytes, sialoadhesin on macro-phages, and myelin-associated glycoprotein on neuronal axon myelin sheaths can show preferences for or lack of binding to Neu5Gc (27). Thus, the genetic defect in humans causing loss of or reduction in Neu5Gc expression could potentially influence a wide variety of biological and pathological processes.
Despite the present finding of a major mutation in the human hydroxylase gene, hu-man fetal tissues (47) and certain human tumors (47–49) have been reported to con-tain Neu5Gc in small amounts. Some humans with malignancies or with certain in-flammatory or infectious diseases are also known to spontaneously develop antibo-dies against Neu5Gc (66-68).We have also noted small traces of an HPLC peak cor-responding to Neu5Gc in some human tissues (37).Taken together,these data indi-cate that the capacity to synthesize Neu5Gc may not be lost completely in humans. Further studies will be needed to ascertain whether this represents the up-regulation of another minor pathway for the synthesis of Neu5Gc, such as the suggested use of the donor glycolyl-CoA (69, 70), or the incorporation of Neu5Gc from dietary sources (71).
In this study, we have isolated a human hydroxylase cDNA that includes the codons corresponding to the initiator methionine of the mouse and chimpanzee hydroxylase. In contrast, the recent study of Irie et al. reported an incomplete cDNA that does not include this 5′ region (50). This 5′-truncated cDNA gave them a protein product in an in vitro translation reaction,leading to the assumption that a methionine codon down- stream of the 92-bp deletion is the primary initiator for translation of a truncated pro-tein representing >80% of the carboxyl end of the hydroxylase.They went on to show that such a truncated protein had no residual hydroxylase activity when expressed in COS cells. However, because we find an intact primary initiator methionine codon corresponding to those of the mouse and chimpanzee proteins, we predict a much shorter truncated polypeptide involving the amino terminus of the protein. Thus, it remains to be seen whether the protein product studied in vitro by Irie et al. (50) actually exists in vivo.
We should also note that there is a difference between the 3′ ends of the major cDNAs reported by the two groups. However, we also isolated an alternative human transcript with identity to the 3′ region reported in their paper, as well as other clones with various deletions and insertions. Although we did not conduct an extensive search in a chimpanzee library, we encountered only one such anomaly in the chim-panzee clones that we isolated. We speculate that the many abnormalities and vari-ations of the 3′ region of the human hydroxylase are reflective of a gene that became nonfunctional in recent evolutionary times, and is undergoing a random degenera-tion.Regardless,if it were possible to determine when the loss of Neu5Gc expression occurred in relation to hominid evolution, this may shed light on the controversial question of the origin of modern humans (72). Unfortunately, autosomal DNA from fossils >30,000 years B.P. is extremely difficult to recover (73).
Finally, given the DNA melting curves and gene sequence data indicating an almost 99% identity of the chimpanzee genome to that of the human (9-16),it has been sug-gested that the genetic changes that result in morphological and functional differen-ces between these species are subtle,e.g., minor changes in the promoter regions of critical genes and/or specific changes in the amino acid sequence of some gene pro-ducts (4). However,the present data emphasize that the genetic differences between humans and the great apes can include major gene alterations.It remains to be seen whether the mutation in CMP-sialic acid hydroxylase can explain some of the drama-tic morphological and functional changes that occurred during the evolution of hu-mans. In this regard, perhaps the most intriguing observation is that no matter what the level of Neu5Gc was in other parts of the body, the amounts in the brain were al-ways very low in a variety of animals studied (34-36), including the chimpanzee (37). This correlates with very low levels of hydroxylase message in the mouse brain (45). These data indicate that, for unknown reasons, Neu5Gc expression is not desirable in the vertebrate brain. Of course, in humans, the last traces of Neu5Gc in the brain are eliminated, because of the genomic mutation in the hydroxylase. It remains to be seen whether this results in any evolutionary advantage for human brain development or function.
Acknowledgments
We thank Peter Parham (Stanford University) for providing the lymphoblastoid lines, Alanna McCall and Huda Zoghbi for the chromosome 6 hybrid panel, and A. Craig Chinault and Barbara Boggs for assistance with FISH studies.This work was suppor- ted by US Public Health Service Grant RO1-GM32373 and the Mathers Foundation. S.T.W is an Investigator of the Howard Hughes Medical Institute.
ABBREVIATIONS
| Neu5Gc | N-glycolylneuraminic acid |
| Neu5Ac | N-acetylneuraminic acid |
| BAC | bacterial artificial chromosome |
| FISH | fluorescence in situ hybridization |
| EST | expressed sequence tag |
| TdT | terminal deoxynucleotidyltransferase |
| EBV | Epstein–Barr virus |
| RACE | rapid amplification of cDNA ends |
| UTR | untranslated region |
Footnotes
References
https://www.nobelprize.org/prizes/medicine/2022/press-release/
" The Nobel Prize in Physiology or Medicine 2022
Press release
English
English (pdf)
Swedish
Swedish (pdf)
![]()
Press release
2022-10-03
The Nobel Assembly at Karolinska Institutet
has today decided to award
the 2022 Nobel Prize in Physiology or Medicine
to
Svante Pääbo
for his discoveries concerning the genomes of extinct hominins and human evolution
Humanity has always been intrigued by its origins. Where do we come from, and how are we related to those who came before us? What makes us, Homo sapiens, different from other hominins?
Through his pioneering research, Svante Pääbo accomplished something seemingly impossible:sequencing the genome of the Neanderthal,an extinct relative of present-day humans. He also made the sensational discovery of a previously unknown homi-nin, Denisova. Importantly, Pääbo also found that gene transfer had occurred from these now extinct hominins to Homo sapiens following the migration out of Africa around 70,000 years ago. This ancient flow of genes to present-day humans has physiological relevance today, for example affecting how our immune system reacts to infections.
Pääbo’s seminal research gave rise to an entirely new scientific discipline; paleogenomics. By revealing genetic differences that distinguish all living humans from extinct hominins, his discoveries provide the basis for exploring what makes us uniquely human.
Where do we come from?
The question of our origin and what makes us unique has engaged humanity since ancient times. Paleontology and archeology are important for studies of human evolution. Research provided evidence that the anatomically modern human, Homo sapiens, first appeared in Africa approximately 300,000 years ago, while our closest known relatives, Neanderthals, developed outside Africa and populated Europe and Western Asia from around 400,000 years until 30,000 years ago, at which point they went extinct.About 70000 years ago,groups of Homo sapiens migrated from Africa to the Middle East and, from there they spread to the rest of the world. Homo sapiens and Neanderthals thus coexisted in large parts of Eurasia for tens of thousands of years. But what do we know about our relationship with the extinct Neanderthals?
Clues might be derived from genomic information. By the end of the 1990’s, almost the entire human genome had been sequenced. This was a considerable accomp-lishment,which allowed subsequent studies of the genetic relationship between diffe-rent human populations. However, studies of the relationship between present-day humans and the extinct Neanderthals would require the sequencing of genomic DNA recovered from archaic specimens.
A seemingly impossible task
Early in his career, Svante Pääbo became fascinated by the possibility of utilizing modern genetic methods to study the DNA of Neanderthals. However, he soon rea-lized the extreme technical challenges, because with time DNA becomes chemically modified and degrades into short fragments. After thousands of years, only trace amounts of DNA are left, and what remains is massively contaminated with DNA from bacteria and contemporary humans (Figure 1). As a postdoctoral student with Allan Wilson, a pioneer in the field of evolutionary biology, Pääbo started to develop methods to study DNA from Neanderthals, an endeavor that lasted several decades.
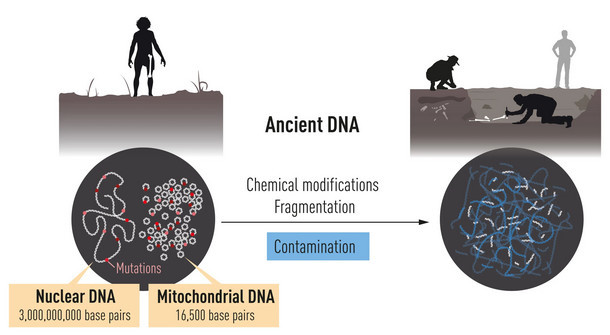
Figure 1. DNA is localized in two different compartments in the cell. Nuclear DNA harbors most of the genetic information, while the much smaller mitochondrial genome is present in thousands of copies. After death, DNA is degraded over time and ultimately only small amounts remain. It also becomes contaminated with DNA from e.g. bacteria and contemporary humans.
In 1990, Pääbo was recruited to University of Munich, where, as a newly appointed Professor, he continued his work on archaic DNA. He decided to analyze DNA from Neanderthal mitochondria – organelles in cells that contain their own DNA. The mitochondrial genome is small and contains only a fraction of the genetic information in the cell, but it is present in thousands of copies, increasing the chance of success. With his refined methods, Pääbo managed to sequence a region of mitochondrial DNA from a 40,000-year-old piece of bone. Thus, for the first time, we had access to a sequence from an extinct relative. Comparisons with contemporary humans and chimpanzees demonstrated that Neanderthals were genetically distinct.
Sequencing the Neanderthal genome
As analyses of the small mitochondrial genome gave only limited information, Pää-bo now took on the enormous challenge of sequencing the Neanderthal nuclear ge-nome. At this time, he was offered the chance to establish a Max Planck Institute in Leipzig, Germany.At the new Institute,Pääbo and his team steadily improved the me-thods to isolate and analyze DNA from archaic bone remains.The research team ex- ploited new technical developments,which made sequencing of DNA highly efficient. Pääbo also engaged several critical collaborators with expertise on population gene-tics and advanced sequence analyses. His efforts were successful. Pääbo accomp-lished the seemingly impossible and could publish the first Neanderthal genome se-quence in 2010. Comparative analyses demonstrated that the most recent common ancestor of Neanderthals and Homo sapiens lived around 800,000 years ago.

Figure 2.A.Pääbo extracted DNA from bone specimens from extinct hominins. He first obtained a bone fragment from Neandertal in Germany,the site that gave name to the Neanderthals.Later,he used a finger bone from the Denisova Cave in southern Siberia, the site that gave name to Deni-sovans. B. Phylogenetic tree showing the evolution and relationship between Homo sapiens and the extinct hominins. The phylogenetic tree also illustrates the gene flows discovered by Pääbo.
Pääbo and his co-workers could now investigate the relationship between Neander-thals and modern-day humans from different parts of the world. Comparative analy-ses showed that DNA sequences from Neanderthals were more similar to sequen-ces from contemporary humans originating from Europe or Asia than to contempora- ry humans originating from Africa.This means that Neanderthals and Homo sapiens interbred during their millennia of coexistence.In modern day humans with European or Asian descent, approximately 1-4% of the genome originates from the Neanderthals (Figure 2).
A sensational discovery: Denisova
In 2008, a 40,000-year-old fragment from a finger bone was discovered in the Deni-sova cave in the southern part of Siberia. The bone contained exceptionally well-pre-served DNA, which Pääbo’s team sequenced. The results caused a sensation: the DNA sequence was unique when compared to all known sequences from Neander-thals and present-day humans. Pääbo had discovered a previously unknown homi-nin,which was given the name Denisova.Comparisons with sequences from contem- porary humans from different parts of the world showed that gene flow had also oc-curred between Denisova and Homo sapiens. This relationship was first seen in po-pulations in Melanesia and other parts of South East Asia,where individuals carry up to 6% Denisova DNA.
Pääbo’s discoveries have generated new understanding of our evolutionary history. At the time when Homo sapiens migrated out of Africa, at least two extinct hominin populations inhabited Eurasia. Neanderthals lived in western Eurasia, whereas Denisovans populated the eastern parts of the continent. During the expansion of Homo sapiens outside Africa and their migration east, they not only encountered and interbred with Neanderthals, but also with Denisovans (Figure 3).
Paleogenomics and its relevance
Through his groundbreaking research, Svante Pääbo established an entirely new scientific discipline, paleogenomics. Following the initial discoveries, his group has completed analyses of several additional genome sequences from extinct hominins. Pääbo’s discoveries have established a unique resource,which is utilized extensively by the scientific community to better understand human evolution and migration.
New powerful methods for sequence analysis indicate that archaic hominins may also have mixed with Homo sapiens in Africa. However,no genomes from extinct ho- minins in Africa have yet been sequenced due to accelerated degradation of archaic DNA in tropical climates.
Thanks to Svante Pääbo’s discoveries, we now understand that archaic gene se-quences from our extinct relatives influence the physiology of present-day humans. One such example is the Denisovan version of the gene EPAS1, which confers an advantage for survival at high altitude and is common among present-day Tibetans. Other examples are Neanderthal genes that affect our immune response to different types of infections.

Figure 3. Pääbo’s discoveries have provided important information on how the world was popula-ted at the time when Homo sapiens migrated out of Africa and spread to the rest of the world. Neanderthals lived in the west and Denisovans in the east on the Eurasian continent. Interbree-ding occurred when Homo sapiens spread across the continent,leaving traces that remain in our DNA.
What makes us uniquely human?
Homo sapiens is characterized by its unique capacity to create complex cultures, advanced innovations and figurative art, as well as by the ability to cross open water and spread to all parts of our planet (Figure 4).Neanderthals also lived in groups and had big brains (Figure 4). They also utilized tools, but these developed very little during hundreds of thousands of years.The genetic differences between Homo sapiens and our closest extinct relatives were unknown until they were identified through Pääbo’s seminal work. Intense ongoing research focuses on analyzing the functional implications of these differences with the ultimate goal of explaining what makes us uniquely human.

Figure 4. Pääbo’s seminal work provides a basis for explaining what makes us uniquely human.
Key publications
Krings M, Stone A, Schmitz RW, Krainitzki H, Stoneking M, Pääbo S. Neandertal DNA sequences and the origin of modern humans. Cell. 1997:90:19-30.
Green RE, Krause J, Briggs AW, Maricic T, Stenzel U, Kircher M, Patterson N, Li H, Zhai W,Fritz MH,Hansen NF,Durand EY, Malaspinas AS, Jensen JD, Marques-Bonet T, Alkan C, Prüfer K, Meyer M, Burbano HA, Good JM, Schultz R, Aximu-Petri A, Butthof A, Höber B, Höffner B, Siegemund M, Weihmann A, Nusbaum C, Lander ES, Russ C, Novod N, Affourtit J, Egholm M, Verna C, Rudan P, Brajkovic D, Kucan Ž, Gušic I, Doronichev VB, Golovanova LV, Lalueza-Fox C, de la Rasilla M, Fortea J, Rosas A, Schmitz RW, Johnson PLF, Eichler EE, Falush D, Birney E, Mullikin JC, Slatkin M, Nielsen R, Kelso J, Lachmann M, Reich D, Pääbo S. A draft sequence of the Neandertal genome. Science. 2010:328:710-722.
Krause J, Fu Q, Good JM, Viola B, Shunkov MV, Derevianko AP, Pääbo S.The com- plete mitochondrial DNA genome of an unknown hominin from southern Siberia. Nature. 2010:464:894-897.
Reich D, Green RE, Kircher M, Krause J, Patterson N, Durand EY, Viola B, Briggs AW, Stenzel U, Johnson PL, Maricic T, Good JM, Marques-Bonet T, Alkan C, Fu Q, Mallick S, Li H, Meyer M, Eichler EE, Stoneking M, Richards M, Talamo S, Shunkov MV, Derevianko AP, Hublin JJ, Kelso J, Slatkin M, Pääbo S. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature. 2010:468:1053-1060.
Meyer M, Kircher M, Gansauge MT, Li H, Racimo F, Mallick S, Schraiber JG, Jay F, Prüfer K, de Filippo C, Sudmant PH, Alkan C, Fu Q, Do R, Rohland N, Tandon A, Siebauer M,Green RE,Bryc K,Briggs AW, Stenzel U, Dabney J, Shendure J, Kitzman J, Hammer MF, Shunkov MV, Derevianko AP, Patterson N, Andrés AM, Eichler EE, Slatkin M, Reich D, Kelso J, Pääbo S. A high-coverage genome sequence from an archaic Denisovan individual. Science. 2012:338:222-226.
Prüfer K,Racimo F,Patterson N,Jay F,Sankararaman S,Sawyer S, Heinze A, Renaud G,Sudmant PH,de Filippo C,Li H,Mallick S,Dannemann M,Fu Q, Kircher M, Kuhlwilm M,Lachmann M,Meyer M, Ongyerth M, Siebauer M, Theunert C, Tandon A, Moorjani P, Pickrell J, Mullikin JC, Vohr SH, Green RE, Hellmann I, Johnson PL, Blanche H, Cann H, Kitzman JO, Shendure J, Eichler EE, Lein ES, Bakken TE, Golovanova LV, Doronichev VB, Shunkov MV, Derevianko AP, Viola B, Slatkin M, Reich D, Kelso J, Pääbo S. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. 2014:505: 43-49.
>Svante Pääbo was born 1955 in Stockholm, Sweden. He defended his PhD thesis in 1986 at Uppsala University and was a postdoctoral fellow at University of Zürich, Switzerland and later at University of California,Berkeley,USA. He became Professor at the University of Munich, Germany in 1990. In 1999 he founded the Max Planck Institute for Evolutionary Anthropology in Leipzig, Germany where he is still active. He also holds a position as adjunct Professor at Okinawa Institute of Science and Technology, Japan.
Illustrations: © The Nobel Committee for Physiology or Medicine. Illustrator: Mattias Karlén
The Nobel Assembly, consisting of 50 professors at Karolinska Institutet, awards the Nobel Prize in Physiology or Medicine. Its Nobel Committee evaluates the nomi- nations. Since 1901 the Nobel Prize has been awarded to scientists who have made the most important discoveries for the benefit of humankind.
Nobel Prize® is the registered trademark of the Nobel Foundation "
Kommentit