Ukrainalainen ”Maidan-HP-tieteilijä”…
Tällaiset asiat jos mitkä, tieteen väärentäminen, osoittavat myös ketkä Ukrainan kriisissä ovat väärässä, väärällä asialla – ja ketkä siis oikeassa…

I have been interested in the physical properties of mental imagery since I was nine years old, and was involved in related research since my undergraduate studies. My current research focuses on the neurological basis of imagery. My work spans the intersection of neuroscience, linguistics, primotology, and paleoanthropology. In the past, I have conducted research in neurophysiology of memory, cardiopulmonary acoustics, and optical vibrometry. I have co-founded several successful companies, received numerous awards from NSF, NIH, and DOD, and directed the development of several FDA approved medical devices.
In 2008,after fifteen years of research,I allowed myself to speculate on the subject of neurobiology of imagination and evolution of language,and published the first edition of “On the Origin of the Human Mind.” The second edition of the book was published in 2014.
I earned my undergraduate degree in astrophysics from St. Petersburg Technical University, under prof.Zhores Alferov,the winner of the 2000 Nobel Prize in physics. I received Master of Science in Biomedical Engineering and my Ph. D. in neuro-science from Boston University.I have been since teaching at Boston University for nearly two decades and have authored over 100 scientific publications, book chapters, and conference presentations. I have also periodically taught Tufts Medical School students. I live in Boston with my wife and two children.
Classes currently taught in Boston University – join me to learn about the brain and the body – I love teaching:
- The Neurobiology of Consciousness and Evolution of Language – MET BI566 (Fall and Spring) and CAS NE594 / BI594 (Summer I)
AV9/4/20191The Neurobiology of Consciousness and Evolution of Language Class description: Your brain is a bizarre device, set in place through natural selection of your ancestors and your own experience. One thing that clearly separates your brain from the brain ofany other non-human animal is the propensity of your brain for ima-gination and creativity. In this class we will dive into the neuroscience of imagination: from neurons to memory to neurological mechanism of generating novel conscious experiences. We willstudy what makes your brain unique and the selectional forces that shaped the brains of our ancestors. We will discuss what makes human langu-age special and how it evolved.This interdisciplinary class is intended for paleoanth-ropologists who want to learn neuroscience, psychologists who are interested in the question of the origin of language, biologists who are interested in the uniqueness of the human mind, neuroscientists who want an exposure to paleoanthropology and linguistics, philosophers fascinated by neurological basis of behavior and other stu-dents interested in an understanding of the mind of a man and the evolution of the brain.
TÄYTTÄ HEWONPASKAA!!!
- Systems Physiology – MET BI 315 (Spring)
- Human Physiology – MET BI 211 (Fall)
- Introduction to Neuroscience – CAS NE101 (N/A)
TED: The neuroscience of imagination
Recent inteviews: Cognitum Episode 1 | Cognitum Episode 2
The interview with Professor Voices
Current Research at ImagiRation
https://www.pirkanblogit.fi/2019/risto-koivula/vaara-teoria-kielen-synnysta/

Kuva:Tämä 37000 vuotta vanha Saksasta löydetty patsas esittää leijonan ja ihmisen yhdistelmää. Jotta taiteilija on voinut luoda tällaisen, hänen on pitänyt ensin kuvitella mielessään ihmisen ja leijonan yhdistel- mä. Taideteos viittaa vahvasti siihen, että ihmiset olivat saavuttaneet etuotsalohkon synteesin 37000 vuotta sitten. Wikimedia Commons
RK: Tekniikan maailma lykkää Wanhaa kumottua hewonpaskaa: kieli esimer-kiksi EI KEHITY GEENIEVOLUUTION mukaan, ja ”Nicaraguan kuurojen lasten keskenään keksimä uusi viittomakieli” oli Noam Avram Chomskyn huijausta.
Uusi teoria selittää, miten ihminen sai kielitaidon – 70000 vuotta sitten ilmestyneen mutaation seurauksena lapset keksivät keskenään nykykielen perusteet
Ihmislaji teki suuren henkisen loikan noin 70000 vuotta sitten, kun ihmiset oppivat puhumaan kieliä ja heille kehittyi nykyisen kaltainen mielikuvitus. Tutkijoiden keskuudessa on kuitenkin pitkään kiistelty sitä, mikä johti tähän loikkaan.
Nyt Bostonin yliopiston neurotieteilijä Andrej Višedskij esittää uudessa tutkimuk-sessaan hypoteesin,joka saattaa selittää kielitaidon kehittymisen ihmiselle. Tutkimus on julkaistu Research Ideas and Outcomes -julkaisussa.
Arkeologiset ja geneettiset tutkimukset ovat pitkään viitanneet vahvasti siihen, että ihmiselle on kehittynyt äänteiden tuottamiseen vaadittavat fysiologiset piirteet jo ennen kuin nykyihmiset erkaantuivat neandertalinihmisistä 600000 vuotta sitten. Ihmiset kykenivät siis 600000 vuotta sitten tuottamaan yhtä paljon samanlaisia ääniä kuin nykyisissä kielissä käytetään.
Siitä huolimatta arkeologiset löydöt viittaavat siihen, että ihmisille on kehittynyt kyky nykyisen kaltaiseen mielikuvitukseen ja symboliikan käyttöön vasta 70000 vuotta sit-ten. Puhe-elinten kehittymisen ja nykyisen kaltaisen puheen ja mielikuvituksen kehit-tymisen välillä on siis puolen miljoonan vuoden pituinen ajanjakso,ja tämä on pitkään hämmentänyt tutkijoita.
Tutkiessaan lasten mielikuvituksen kehittymistä Višedskij on kollegoineen havainnut, että lasten mielikuvituksen tietyn komponentin kehi- tyksessä on tärkeä aikaikkuna. Jos lapset eivät varhaislapsuudessa altistu kielenkäytölle, he eivät koskaan saavuta sellaista mielikuvituksen tasoa, jolla voidaan rinnastaa erilaisia käsitteitä. Tätä taitoa kutsutaan etuotsalohkon synteesiksi (Profrontal Synthesis, PFS).
Esimerkki tällaisesta taidosta liittyy Višedskijn mukaan lauseeseen:”Käärme, joka on mäen takana olevan korkean puun vasemmalla puolella olevalla kivellä”. Tällaisen lauseen ymmärtäminen edellyttää PFS:ää, jotta sen kuulija pystyy mielessään yhdistämään sanat yhtenäiseksi kuvaksi.
Kaikki nykykielet sisältävät tällaisia rekursiivisia piirteitä, joissa asioita voidaan yhdistellä eri tavalla ja toistaa loputtomasti. Siksi kielitieteilijät kutsuvat nykykieliä rekursiivisiksi kieliksi.
Vaikka kielioppia ja sanastoa voi oppia aikuisenakin, PFS:ää ei voi oppia aikuisena, jos lapsena ei ole altistunut rekursiivisen kielen kuuntelemiselle.Samalla tavalla mui-naiset ihmiset eivät olisi voineet oppia rekursiivista kieltä aikuisina ja siten pystyneet opettamaan sitä lapsilleen.
Nykylapset pystyvät oppimaan PFS-taidon viiteen ikävuoteen mennessä, ja sen jälkeen se ei ole mahdollista. Esimoderneilla lapsilla tuo aikaikkuna loppui jo kahden ikävuoden kohdalla.
Vyshedskiyn kehittämä matemaattinen malli ennustaa,että jotta kielitaito voi kehittyä, ihmisten piti ylittää nämä evoluutioesteet muutaman sukupolven aikana. Mutaatio, jonka avulla PFS-taitojen kehittyminen on mahdollista viiteen ikävuoteen saakka, on kaikissa nykyihmisissä,mutta sitä ei ole neandertalinihmisissä.Mutaatio myös häviäi-si populaatioissa, jotka eivät käytä rekursiivista kieltä. Malli viittaa siihen, että mutaa-tion ilmestyminen ihmisiin noin 70000 vuotta sitten mahdollisti rekursiivisen kielen syntyminen.
Malli edellyttää, että kaksi tai useampi lapsi saa tämän ”PFS-viivemutaation” ja että nämä lapset viettävät paljon aikaa puhuen keskenään. Mallin mukaan he keksisivät keskenään kielen rekursiivisen kielen elementit. He siirtäisivät mutaation lapsilleen ja opettaisivat rekursiivisen kielen heillekin.
Voi kuulostaa vaikealta uskoa, että lapset keksisivät keskenään rekursiivisen kielen elementit. Tiedote kuitenkin muistuttaa, että samanlaista kielen keksimistä on havaittu esimerkiksi Nicaraguassa kuurojen lasten keskuudessa.
RK: Täyttä huijasta, moneen kertaan paljastettua.
Tästä on kinata päristelty kovasti 20 vuotta sitten.Nuo tuolloin jo armotta kumotutu teoriat olivat silloin ”toisella disinformaatiokierroksella”, jonka syy oli allekirjoittaneelle tuota keskustelua käydessä vielä tuntemoton ”peilineurooniteoria”…
TM: ”PFS:n ja rekursiivisen kielen keksiminen 70 000 vuotta sitten johti siihen, että syntyi käytökseltään uudenlainen laji: ensimmäinen behavioristisesti moderni Homo sapiens”, Višedskij sanoo tiedotteessa.
Tämä mahdollisti sen, että ihmiset pystyivät viestimään asioita toisilleen ennenäke-mättömän tehokkaasti, ja ihmisistä tuli nopeasti maapallon dominoiva eläinlaji.
Ihmiset saivat muun muassa kyvyn saada ansaan suuria eläimiä,mikä johti ravinnon- saannin paranemiseen. Kun populaatio kasvoi nopeasti, ihmiset levisivät nopeasti pois Afrikasta ja asuttivat suuren osan planeetasta.
Esimerkiksi Australiaan nykyihmiset saapuivat 50 000 vuotta sitten. He olivat hyvin paljon nykyihmisen kaltaisia, koska he käyttivät rekursiivista kieltä, jonka PFS-viivemutaatio mahdollisti.
Lue myös
***
Yhdysvaltalaisen Massachusetts Institute of Technologyn (MIT) yleisen kielitieteen emeritusprofessori Noam Chomsky (s.1928) on teoriallaan ns. geneerisestä univer- saalikieliopista” (ainakin ollut) maailman eniten siteerattu tiedekirjoittaja, ainakin jos otetaan huomioon riittävän matalatasoisetkin tiedejulkaisut, osittain johtuen alasta, jolla paljon julkaistaan,osittain muista syistä. Geneerinen on Chomskyn itsensä käyt- tämä termi,joka tarkoittaa geeniperäistä.Hänen teoriansa mukaan kaikkien maailman kielten kieliopilla ja vieläpä sanastollakin on yhteinen aivoperäinen ”syvärakenne”, jonka ilmenemismuotoja ovat konkreettiset kieliopit ja vieläpä sanastotkin.
***
Marx oli oikesaa Feurbach-teesissään, samoin Stalin teoksessaa ”Maxismin kysymyksiä kielitieteessä.
Marxin saavutus on sitäkin merkittävämpi, koska hänellä ei ollut erityistieteellistä kokeellista mittaustietoa kuten esimerkiksi Joosulla.
keskiviikko, 17. elokuu 2011
Marxismi ja frankfurtismi: Feuerbach ei ollutkaan feuerbachilainen?
Yleisesti katsotaan, että Ludwig Feuerbach olisi keksinyt ns. ”ihmisluonnon” käsit- teen, josta hänen ja Hegelin oppia luovasti yhdistellyt Marx sanoutui irti tuotantonsa tunnetuimpiin ydinkohtiin kuuluvissa Feuerbach-teeseissään.
Marxin omalta osaltaan kumoama oppi olisi sitten jäänyt elämään hiljattain Norjassa julkisuutta saaneen FRANKFURTISMIN opin muodossa, jossa (luokka)yhteiskunta ”kiristää ja puristaa” sen olettamaa ”oikeata ihmisluontoa” (sen sijaan, että ”ilmentäisi sellaista”,kuten taas muutoin samaan aksioomaan nojaavilla ns.”sosiobiologisteilla”). Frankfurtismin rajaukseni suhteessa ”sosiobiologiaan”,josta Sovjetskaja ei sano juuri mitään, perustan erityisesti Erich Frommin ideologialle.
Psykologi Jussi Silvonen, joka on tutkinut erityisesti psykologi ja kasvatustietelijä L. S. Vygotskin tuotantoa ja roolia, esittää asiasta uusia näkökohtia, jotka tuovat nuo eri opit hyvin esiin. Silvonen katsoo Marxin ja Feuerbachin olleen ajattelijoina lähempä-nä toisiaan kuin dialektisessa materialismissa on katsottu.Hänen todisteensa osoitta-vat kuitenkin pikemminkin,että ”Ludwig Feuerbach ei ollutkaan feuerbachilainen”, kuin että ”Marx ei olisi ollut marxilainen”… Nimi Feuerbach ei ole toki aatehistoriasta millään tavoin ”unohtumaan päin”, päin vastoin…
[ www.kolumbus.fi ]
JS: ” ”Todellisuudessaan se (ihmisen persoonallisuus) on yhteiskunnallisten suhteiden näyttämö”.
Feuerbach-teesit ja kulttuurihistoriallinen psykologia ”
Jussi Silvonen.
Esitelmä 1) Marxin Feuerbach-teesit tänään seminaarissa Tampereen yliopistolla 16.12.2005”
Hämeemmiäs: ” Otsikko on heti väärin: Marxin Feuerbach-teesien mukaan ”ihmisen ’persoonallisuus’ on yhteiskunnallisten suhteiden (asiaintilojen ym.) ENSEMBLE” eli ”kooste, yhteisesitys”,”(taiteellinen enemmän kuin tieteellinen) kokonaisuus”. (Tämä sama ei tarkoita myöskään samaa kuin dialektiikan kokonaisuus (whole),joka on osa (part, dialektinen vastakohta.) ”Näyttämö” olisi aivan eri asiaa,joka viittaisi sellaiseen, että se olisi jotakin, MISSÄ tapahtuu, ja jolla ”raamilla” olisi oma olennainen ”raami-vaikutuksena” kaikkeen,mitä ”näyttämöllä” tapahtuu.Tuollaista esittää Steven Pinker, mutta ei Karl Marx.
***
http://imagiration.com/a3/2014_Vyshedskiy_OnTheOriginOfTheHumanMind_Edition2.pdf
" Praise for On The Origin Of The Human Mind
“I like the idea of mental synthesis very much ... I quite agree that language evol-ved in a way that facilitates synthesis and transmission of the synthesized mental image. ... I don't think there can be much doubt,purely conceptually,that language was a late arrival.Whatever mutation provided the key to it would have had no se- lectional advantage at all, and would have just been a useless “organ,” if it could not have linked up to pre-existing thought systems.”
— NOAM CHOMSKY, Professor Emeritus of Linguistics, MIT
“Boston University’s Andrey Vyshedskiy brings a neuroscientist’s perspective to the discussion of human mental history in On the Origin of the Human Mind.”
—Scientific American Mind (July 2009)
“I found the Mental Synthesis theory stimulating and provocative. The author puts forward an explanation for the evolution of the human mind based on predator detection that led to increased visual mental analysis which set the stage for visual mental syntheses. The author presents an impressive array of recent re-search on the brain with up to date references that are highly relevant to his case and the origin of mind. For me, the most interesting aspect of the book centers on the significance of imagination for understanding the evolution of the mind, which, as the author states, has not been given enough attention in academic circles. This book brings forth a great deal of interesting new research that would be of interest both to the informed reader and the general public.”
— DEREK HODGSON, Professor of Cognitive Archaeology, University of York
“I strongly recommend this book to everyone interested in the human evolution, primarily on the strength of
(1) the author’s original thesis that detecting hidden motionless predators was a driving force for the evolution of mental synthesis, and
(2) the author’s overall coherency and clarity in integrating a sufficiently wide range of evidence for his mental synthesis theory (such as research on language, vision science, neuroanatomy,evolutionary history of primates,and archaeological / paleontological evidence of tool and art creation and usages).”
— RICHARD J. HARRINGTON, Professor of Anthropology, Integrative Centers for Science and Medicine
ii
“It is with great enthusiasm I recommend this book to everyone interested in the study of the human mind. The theory of integration of neuronal ensembles allowing for a uniquely human experience of “mental synthesis” is fascinating and is presented in a clear and easy-to-understand language. Author’s diverse back-ground in neuroscience, bioengineering, and humanities allowed him to integrate all these fields into a coherent and fascinating text. His contribution to our com-mon knowledge of evolutionary neurobiology will be extremely valuable to estab-lished neuroscientists, medical and graduate students and interested people of all professions alike.”
— MARIA K. HOUTCHENS, M.D., MMSc, Professor of Neurology, Harvard Medical School
“I read the book in one sitting, which was an easy task owing to the author’s concise writing and knack of explaining science with simple clarity.”
— NICHOLAS J MULCAHY, Hon Research Fellow, Univ. of Queensland, Australia
“I enjoyed a lot reading Vyshedskiy’s book. I’m impressed by the quality of the book on its main topic: qualitative differences between human and animal brains and the role of ‘mental synthesis’. The book devotes a lot of pages to the impor-tance of neural synchronisation. This is a strong point to my view, as I share with the author the idea that (group/phase) synchronisation plays a major role in fea-ture binding and consciousness. The book makes also a strong point about men- tal synthesis. I made a similar point (though with less emphasis) in Why we talk (2007; 2000, French version). The book is very didactic and well-documented on various topics, especially brain functional anatomy. It is also excellent on evolutio-nary facts. I learned from it, and I will certainly recommend it to my students at least for this reason. One can only admire the author for the breadth of his knowledge and the clarity of his account.”
— JEAN-LOUIS DESSALLES, Professor at Telecom ParisTech, author of Why We Talk: The Evolutionary Origins of Language
“I like this book very much. Like the first edition, it is engagingly written and beau-tifully illustrated. I have to say that I am not entirely convinced that mental synthe-sis is what distinguishes the human mind,but the author does make a good case.”
— MICHAEL CORBALLIS, Professor Emeritus of Psychology, University of Auckland, author of The Recursive Mind

Published by MobileReference
Copyright © 2014 Dr. Andrey Vyshedskiy
All rights reserved.
No part of this book may be reproduced, stored in a retrieval system, or transmitted by any means without the written permission of the author except for brief quotations embodied in critical articles and reviews. Second edition, 2014
The first edition was published in 2008 by MobileReference.
ISBN-13: 978-1492963615
ISBN-10: 1492963615
Audience
The book speaks best to readers who want to approach the mind from a scientific perspective. The book is written in easy-to-read engaging style.
No previous knowledge in psychology, paleoanthropology, or neuroscience is necessary.
Front cover
“Lion-man” statuette carved out of mammouth-tusk, H 296 mm
Site: Hohlenstein-Stadel-cave, Asselfingen, Baden-Württemberg,
Germany
Upper Paleolithic period (Aurignacien), approx. 35 000 BC
Inv. Ulmer Museum Prä Slg. Wetzel Ho-St. 39/88.1
Photo Thomas Stephan © Ulmer Museum, Ulm, Germany
Please visit the book’s website to watch videos, read supplemental materials, or to contact the publisher:
www.MobileReference.com/mind
Part I. Neuroscience of imagination
iii
“I thoroughly enjoyed reading the book. I think the idea about “mental synthesis” is brilliant and that it should enter the literature as an alternative to the other theories that explain the origin of humans.”
— FREDERICK E WASSERMAN, Professor of Biology, Boston University
“The value of Vyshedskiy's book may rest on his highly original concept of mental
synthesis and its possible neural basis.I am not a neuroscientist and cannot judge the plausibility of the neural mechanisms proposed by the author for linking sen- sory areas of the brain with the prefrontal cortex. But he marshals an impressive body of empirical evidence and develops a persuasive argument. If this work holds up to neuroscientific scrutiny, it will prove a valuable contribution to an account of the neural basis for displaced reference, and a nice complement to Derek Bickerton's recent work, "More than Nature Needs" (Harvard, 2014).”
— MICHAEL STUDDERT-KENNEDY, Professor Emeritus of Psychology, Univer-sity of Connecticut, Professor Emeritus of Linguistics at Yale University, co-author of Approaches to the Evolution of Language
“This is a book with a sweeping scope and a grand vision. It incorporates insights from neuroscience,linguistics and paleoanthropology, which can only be achieved by someone with rigorous scientific training who is at the same time courageous enough to cross many boundaries between academic discipline.
— JEN-WEI LIN, Professor of Neuroscience, Boston University
“The book is well written, and serves to stimulate consideration of what factors contributed to the emergence of ‘behaviourally modern’ Homo sapiens.“
— FRANCIS THACKERAY, Professor of Anthropology, University of the Witwatersrand
“I had an opportunity and pleasure to read one of the first copies of this exciting
book. This is one of the few successful attempts to present a coherent model of
the human mind in an engaging and fascinating manner. I’ve enjoyed the discus- sion of numerous animal and human intelligence experiments, as well as compre- hensive analysis of the development of the visual system on the evolutionary timescale. I found the neurobiological explanation of visual object representation in human minds convincing. The book is a captivating read that ultimately leads to a well thought out theory of what makes us humans.”
— ALEX GANELIS
iv
“... the theory of the evolution of “mental synthesis” through a refinement of the
visual system and the theory of conscious thought the voluntary synchronization
of neuronal ensembles were enlightening for me because they showed that a
credible scientific explanation of these two facts is possible. ... I do believe now
that they are within the reach of current science. Previously, these two fields were as enigmatic to me as a light bulb might have been to an Australopithecus.”
“... The book presents a very logical theory of a human mind. This logic is easy to
understand and follow. I recommend this book to anyone who is curious, who
always wants to get to the bottom of things. The book is a lot of fun to read.”
“ On the Origin of the Human Mind is a highly important work because it sheds
light on that most defining, yet elusive, quality of our nature -- the imagination. I
agree with Einstein's view that imagination is more important than knowledge. I
have believed this for many years, even before really being able to articulate why. Dr. Vyshedskiy's book has been a great asset to me (as well as a pleasure) be-cause it provides a more concrete explanation for what most people only know by intuition - namely, that the human mind is unique. By bringing together the evi-dence from both our species' evolutionary history as well as the latest in neurolo- gical research, I think he makes a highly compelling case for the theory of mental synthesis. This is one of those rare books that I wish I could get everybody to read!”
— DANE WO

1
Introduction
“γνῶθι σεαυτόν”
(“know thyself”)
Inscribed in the forecourt of the Temple of Apollo at Delphi
While studying the neuroscience of consciousness, I was struck with certain facts about mental imagery that seemed to shed some light on the process of the evo-lution of the human mind. The origin of the human mind remains one of the greatest mysteries of all times. The last 150 years, since Charles Darwin proposed that spe-cies evolve under the influence of natural selection (Darwin C,1859),have been mar- ked by great discoveries. Molecular biology described the genetic principles under-lying species evolution and identified specific changes in the human genome since our lineage split off from the chimpanzee line about six million years ago (Somel M, 2013). Great paleontological discoveries have filled that span of six million years of human evolution with a number of intermediate species that display both human- and ape-like characteristics. However, the discussion of the evolution of the human intellect and specific forces that shaped the underlying brain evolution is as vigorous today as it was in Darwin’s times.
At the center of the predicament about the origin of the human mind lies the question of human uniqueness. Most scientists agree that humans possess a unique intellect that sets us apart from other animals (for ex. see Gazzaniga MS, 2008). However when any individual skill is considered, researchers invariably point to a comparable skill among non-human primates. Scientists used to think that only humans made and used tools. Sherwood Washburn, the great American physical anthropologist, has even suggested that the use of tools was the main driving force of human evolu-tion. He wrote, “It was the success of the simplest tools that started the whole trend of human evolution and led to the civilizations of today” (Washburn S, 1960).
2
Washburn felt that tools were responsible for the changes in hominid teeth, hands, brain, and pelvises; tools,in effect,changed the pressures of natural selection and thus formed the man.However since the late 1960s, researchers have found numerous examples of animals using tools in the wild (Goodall,1968; McGrew,1992; Whiten, 1999; Pruetz J,2007), which pretty much discredited the hypothesis of intellect acquisition through tool use. Scientists used to think that only humans could have an expanded vocabulary, but it is now known that chimpanzees, bonobos, and gorillas can be taught to use hundreds of words meaningfully (Terrace HS, 1979; Patterson FG, 1978; Savage-Rumbaugh ES, 1994). Scientists used to think that only humans could count, but it has been dis-covered that chimpanzees have arithmetical skills (Inoue S, 2007). "
HM: This is a lie. No animal can count numerically.
https://hameemmias.vuodatus.net/lue/2017/01/osasiko-ihmetrolli-harmaapapukaija-alex-2007-oikeasti-laskea
A.V.: " Many social functions,such as altruism,understanding another per- son’s cognitive state [a lie, HM], social cooperation, and cultural transmis-sion, which were once thought to be human-specific, have recently been described in various forms in chimpanzees,bonobos and other great apes (Hare B,2010; Luncz LV,2012; Whiten A,2009; Horner V, 2011
Call J, [Tomasello, M, 2008; ... "
[HM:This competent and creditable research of Josep Call and of (a FORMER "mirror neuron" pseudoscientisti) Michael Tiomasello proves that chimpanzees DON´T HAVE any SHARED INTENTION which is uniquely human and an absolute prerequisition on human thinking and consciousness!
https://www.eva.mpg.de/documents/Elsevier/Call_Does_TrendsCogSci_2008_1554401.pdf
Does the chimpanzee have a theory of mind? 30 years later
Josep Call and Michael Tomasello
Max Planck Institute for Evolutionary Anthropology, Deutscher Platz 6, D-04103 Leipzig, Germany
On the 30th anniversary of Premack and Woodruff’s seminal paper asking whether chimpanzees have a theory of mind, we review recent evidence that suggests in many respects they do, whereas in other respects they might not. Specifically, there is solid evidence from several different experimental paradigms that chimpanzees understand the goals and intentions of others, as well as the perception and know-ledge of others. Nevertheless, despite several seemingly valid attempts, there is currently no evidence that chimpanzees understand false beliefs. Our conclusion for the moment is, thus, that chimpanzees understand others in terms of a perception - goal psychology, as opposed to a full-fledged, human-like belief–desire psychology.
Introduction
In 1978 Premack and Woodruff asked, ‘Does the chimpanzee have a theory of mind?’ In this brief review we attempt to answer this question based on much research that has been con-ducted in the 30 years since that time, particularly in the last decade or so (see Tomasello and Call [1] for the state of the art as of the mid-1990s). The answer will not be a simple yes or no, however, because part of the progress that has been made in recent years is a recognition that there are many different ways in which organisms might understand the psychological functioning of others.
... In a broad construal of the phrase ‘theory of mind’,then, the answer to Premack and Wood-ruff’s pregnant question of 30 years ago is a definite yes, chimpanzees do have a theory of mind. But chimpanzees probably do not understand others in terms of a fully human-like belief – desire psychology in which they appreciate that others have mental repre-sentations of the world that drive their actions even when those do not correspond to reality.
And so in a more narrow definition of theory of mind as an understanding of false beliefs,
the answer to Premack and Woodruff’s question might be no, they do not.
Why chimpanzees do not seem to understand false beliefs in particular – or if there might be some situations in which they do understand false beliefs – are topics of ongoing research.
... "
Vyshedskiy does not understand what he reads and links.
A.V.:"These findings cast doubts on the “social brain hypothesis", which argues that human intelligence evolved primarily as a means of surviving and reproducing in large and complex social groups (Dunbar RIM, 1998). At the start of the 21st century, there is still no consensus as to what makes the human intellect unique, and without an understanding of human uniqueness, it is very difficult to meaningfully discuss either the genetic evolution of the human brain, or the relevant changes in brain morphology, or even the driving forces behind the evolution of human intelligence.
Over two decades ago, when the question of human uniqueness was first presented to me by a colleague, it occurred to me that I should look for the difference between humans and other animals in respect to mental imagery. I have been interested in the physical properties of mental imagery since I was nine years old, and was involved in related research since my undergraduate studies. Having been trained in neuroscience, I set out to understand the neurological basis of imagery pertaining to the differences between humans and other animals. In 2008, after fifteen years of research, I allowed myself to speculate on the 3 subject, and published the first edition of “On the Origin of the Human Mind.”

Some of the most time-honored questions in philosophy, psychology, and neurosci-ence center on the uniqueness of the human mind. How do we think? What makes us so different from all the other animals on planet Earth? What was the process that created the human mind? Is this process unique or can it be repeated on other pla-nets? The book "On the Origin of the Human Mind" attempts to provide an answer to these questions. It is organized into three chapters:
Chapter I "Uniqueness of the Human Mind" introduces the reader to recent research into animal behavior, communication, culture and learning, as well as controlled ani-mal intelligence experiments and offers a new hypothesis of what makes the human mind unique.
Chapter II "Evolution of the Human Mind" combines latest genetics research and archeological discoveries to help readers understand hominid evolution. The author discusses the forces that influenced the development of the hominid intelligence and offers a step-by-step theory that links improvement in visual information processing to speech development and to the types of stone tools manufactured by the hominids.
Chapter III "The Neurological Basis of Conscious Experience" takes the reader on an exciting journey into the neurobiology of the human mind. The author introduces the reader to the structure and function of the brain and then presents recent insights into brain organization derived from cognitive psychology, brain imaging, animal experiments, and the studies of patients with diseases of the brain. The book concludes with a unifying theory of the mind and a discussion of the evolution of the human brain and the uniqueness of the human mind from the neurological perspective.
Audience
The book speaks best to readers who want to approach the mind from a scientific perspective. The book is written in easy-to-read engaging style. No previous knowledge in psychology, paleoanthropology, or neuroscience is necessary. "
From that period to the present day,I have continued to work on the same subject. I hope that I may be excused for including this personal infor- mation about the time-frame of my research as I provide it to show that I have not been cursory in coming to my conclusions.
Henri Poincaré, a French mathematician and a philosopher of science, wrote, “Science is built with facts, as a house is built with stones; but a collection of facts is no more a science than a pile of stones is a house. ... Above all, the scientist must make predictions” (Poincaré,1902, translated by Striedter GF,2005). Paleontology, molecular biology, and neuroscience have provided a great number of “facts” concerning human evolution.
This book uses that scientific data to conjecture a thesis on the origin of the human mind. The proposed model connects the dots between the ar-cheological and genetic findings,explains the evolution of stone tools, lan- guage, and culture and yields testable, often counter-intuitive predictions for the neurobiological differences between humans and other primates.
I will start this book by sketching out a neurobiological model of what hap-pens in the human mind when we imagine something that we have never seen before,such as an apple on the back of a whale. This process involves the syntheses of two existing mental images into a new one.
Therefore I will refer to this process as mental synthesis.
HM: It is a wrong "synthesis".
4
Part 1. Neuroscience of imagination
Chapter 1: Object encoding in the brain
... how can a brain perform difficult tasks in one hundred steps that the largest pa-rallel computer imaginable can't solve in a million or a billion steps? The answer is the brain doesn't "compute" the answers to problems; it retrieves the answers from memory. In essence, the answers were stored in memory a long time ago. It only takes a few steps to retrieve something from memory. Slow neurons are not only fast enough to do this,but they constitute the memory themselves. The entire cortex is a memory system. It isn't a computer at all.
JEFF HAWKINS, On intelligence (2004)
1.1 Single neuron recording
One of the most exciting experiments that delves into the neuroscience of mental imagery was conducted at the turn of the millennium by Gabriel Kreiman, Christof Koch, and Itzhak Fried at UCLA and Caltech (Krei-man, 2000). The researchers were involved in identifying areas of the brain responsible for abnormal electrical activity in patients with intrac-table epilepsy. To achieve this goal, surgeons implanted electrodes inside the patient’s brain,in and around the hippocampus. The hippocampus is a group of specialized neurons located deep inside the temporal lobe es-sential for forming long-term memories of people, places and events. The electrodes remained in the patient’s brain for approximately a week, and were used to monitor neuronal activity.During this time,the scientists were able to interact with the patients by having them go through a number of specific tasks.This allowed the researchers to directly observe the activity of these patients’ neurons.
The study, which recorded from hundreds of neurons, found that in the majority of cases, neurons that were activated during vision of a particular object were again activated during the recall of the same object. The pa-tients were shown objects from nine categories, one at a time, and their neuronal activity was recorded.
7
The researchers were able to find 49 neurons that were activated during vision, most of which (44) responded selectively to only one of the nine objects. Later, the patients were prompted to recall each one of the ob-jects with their eyes closed. This time researchers were able to pinpoint 33 neurons that fired during visual imagery, 23 of which were selective to exactly one of the recalled objects. Of the 16 neurons that fired during both vision and visual imagery, 14 neurons fired selectively during vision and recall of the SAME object.
For example, a single neuron in the entorhinal cortex of one of the pa-tients showed an increased firing rate when the patient was shown a pic-ture of a baseball, and not when the patient was shown any other object (such as an emotional face or a food item). Later, the same patient was asked to recall each one of the objects, one at a time, and the same neu-ron responded with increased firing only when the patient was recalling the baseball. In another patient, a neuron in the left amygdala increased its firing rate exclusively when the patient viewed a picture of an animal and then again when the patient (now with eyes closed) was prompted to mentally recall the same animal.
The findings of this remarkable study imply that the mechanisms in the human brain that encode objects may be the same as the mechanisms that allow us to recall that object. In other words, this experiment supports the notion that your brain is acting in a very similar way when you use your eyes to recognize an object in an image as when you close your eyes and imagine the object in your “mind’s eye.” A number of other experiments that also support this hypothesis will be discussed later on, but first let us look more closely at exactly how the neurons in our brain encode an object.1
1 Understanding the evolution of the human mind is impossible without knowing the fascinating and complex way in which our brain is organized. If you haven’t studied neuroscience before, you can benefit from taking a look at Appendix 2 which will get you acquainted with some of the basic neuroscience topics essential for the purposes of our discussion.
8

Figure 1.1 A simplified drawing of a typical neuron found in the brain.
The human brain contains more than 100 billion neurons and each neuron can be linked to as many as 10,000 other neurons. All neurons are small cells with a cell body that is about 0.02 millimeters wide (that’s just a bit thinner than an average human hair) and long branch-like projections that can be up to one meter in length. These long projections allow neurons to exchange information over long distances. Neurons exchange information by firing action potentials, which are short 1 millisecond (0.001 seconds) spikes in voltage from a resting membrane potential of negative 70 millivolt (0.070 Volt) to positive 40 milli-volt. Some neurons fire action potentials as often as 100 times per second, while others can go minutes without firing. These spikes in voltage can be measured by electrodes implanted in the brain.

Figure 1.2 Functional organization of the cerebral cortex.
Neurons in the cerebral cortex are organized territorially based on their function. Motor neurons that control muscle movement are located in the motor cortex. Neurons sensitive to touch are located in the somatosensory cortex. Neurons responsible for language com-prehension are concentrated in Wernicke’s area. Neurons responsible for language pro-duction are located in Broca’s area. If you think of a brain as a company, then functional cortical areas correspond to different departments. Each department minds its own business.
10

Figure 1.3 Organization of the visual system. The visual system, which makes up about half of the entire cortex (Barton, 1998), also consists of multiple departments. The primary visual cortex, V1,is located in the occipital lobe. V1 receives information from the retina via the lateral geniculate nucleus in the thalamus. The primary visual cortex is the first cortical area that receives visual information. There is a specialization hierarchy along the visual information pathways. The departments become more specialized the farther the informa- tion moves along the visual pathway. Neurons in the lateral geniculate nuclei can be acti-vated by visual stimulation from either one eye or the other but not both eyes. They res-pond to any change in activity of the retinal neuron that they are connected to. Neurons in V1 can usually be activated by either eye.Neurons in V1 are sensitive to specific attributes such as the orientation of line segments,color,and binocular disparity. Visual information is transmitted from V1 to cortical areas with greater specificity. Neurons in V4 respond selec-tively to aspects of visual stimuli critical to shape identification. Neurons in the inferior tem-poral lobe may respond only when an entire object (such as a face) is present within the visual field.The perception of motion is the major function of the visual area V5,also known as visual area MT
1.2 Objects are encoded by ensembles of neurons
Our visual world appears to consist of meaningful, unified, and stable objects that move in spatially and temporally predictable ways. Similarly when we close our eyes and imagine a visual scene, that scene is also populated by unified and stable objects. “Objects,” therefore, constitute the functional units of perception. Finding object-selective neurons is a great first step, but a full understanding of object encoding requires consi-dering all the neurons in the brain that represent a particular object, not just the single neuron recorded by scientists. Few neuroscientists believe that objects are encoded by single neurons. The so-called “grandmother cell” theory, which asserts that every object (such as your grandmother, hence the name of the theory) is represented by a single cell, was discre-dited in its pure form. Rather, most neuroscientists think that the percep-tion of each object is associated with firing of tens of thousands of neu-rons located throughout the brain (for review see Quiroga, 2008; Buzsáki G, 2004). In the Kreiman experiment, for example, the “baseball neuron” would have been only one of thousands of neurons that fired when the patient perceived a baseball; it just happened to be the only one that was being recorded during the experiment.
These thousands of neurons encode the various characteristics of the baseball, such as its shape, color, texture,etc. (Quiroga & Kreiman, 2009; Waydo et al.,2006). The majority of the neurons activated by the baseball image are located in the primary visual area (usually called V1), which is located in the back of the brain. A smaller number of neurons are activa-ted in the extrastriate areas such as V2 and V4, and an even smaller
number of neurons are activated in the temporal lobe where scientists placed their recording electrodes.
This neuronal arrangement is best represented by a pyramid. The base of the pyramid represents the high number of neurons located in the primary visual area (V1) that encode the contours of the object, its color, spatial depth (by means of binocular disparity), etc. These neurons at the base of the pyramid are relatively non-selective; they are involved in the encoding of a great number of different objects.The middle of the pyramid represents the fewer number of relatively more selective cells in the ex-trastriate areas such as V2 and V4 that encode simple shapes. The top of the pyramid represents an even fewer number of very selective neurons in the temporal lobe. The firing of these selective neurons in the temporal lobe is often associated with a single face, object, or event. For example, the baseball neuron recorded in Kreiman’s experiment was one of these selective neurons. It responded to an image of a baseball but not to any other images presented to the patient.

Figure 1.4 Neuronal ensembles encode objects and events in memory. This illustration shows a schematic representation of a neuronal ensemble overlaid over the ventral visual path. A neuro-nal ensemble pyramid includes a high number of relatively non-selective neurons in V1 (at the base of the pyramid) that encode the contours of the object, its color, the spatial depth, etc.; a fewer number of relatively more selective cells in the extrastriate areas such as V2 and V4 that encode simple shapes; and an even fewer number of very selective neurons in the temporal lobe (the top of the pyramid). It is important to understand that a significant number of selective neu-rons in the temporal lobe are associated with any single neuronal ensemble.Neuronal ensembles include neurons located throughout the cerebral cortex, not limited to V1, V2, V4, and the tempo-ral lobe, but this simplified representation makes it easier to conceptualize a neuronal ensemble. The layers of the pyramid are connected by axons going in both directions that enable the neurons of the ensemble to activate into a common resonant mode.
Part 1. Neuroscience of imagination 12
In addition to the baseball neuron, researchers recording the activity of single neurons in the medial temporal lobe of patients have identified neurons selectively activated by animals (such as spiders and seals), people (such as Bill Clinton, Halle Berry and Jennifer Anniston) and pla-ces (such as the Sydney Opera House and the Tower of Pisa) as well as hundreds of other objects (Quiroga,2005; Quiroga, 2008; Quiroga, 2009). These neurons were activated by objects in an invariant, stable, and ex-plicit manner. This means that the object’s neuron was activated when-ever the patient was shown any image of the particular object even if it displayed the object in a novel position, from another angle or in a diffe-rent context. For example, the Bill Clinton neuron would react strongly whenever Bill Clinton was shown, be it from the front or from the side, whether it was his whole body or just his face, and even when he was in the company of other dignitaries. Remarkably, that same neuron would be activated even when the patient saw the written name of the object, heard the spoken name of the object or even simply recalled the object from memory. That same neuron would stay silent when the patient was exposed to photographs, written or spoken names of other objects.
There is significant scientific evidence that all neurons encoding an ob-ject, such as a baseball, activate synchronously when a person perceives that object (discussed below). Tens of thousands of neurons in various parts of the brain fire one or more action potentials in synchrony. For this reason, neuroscientists call this collection of neurons a neuronal ensemble or neuronal assembly.2
2 At this time,neuronal ensemble and neuronal assembly are used interchangeab-ly. An ensemble has a somewhat more permanent connotation (think of an en-semble of musicians playing a melody in synchrony) while assembly has a more transient meaning (think of a citizen assembly for a single vote). In this mono-graph, I use the term neuronal ensembles to connote more permanent self-orga-nizing structures. A neuronal assembly can become a neuronal ensemble if cells of the assembly repeatedly fire together.
Donald Hebb first developed the theoretical concept of neuronal en-sembles in 1949. He defined “cell assembly” as “a diffuse structure com-prising cells in the cortex and diencephalon, capable of acting briefly as a closed system,delivering facilitation to other such systems” (Hebb, 1949). Hebb suggested that, depending on functional requirements, individual neurons could participate in different neuronal ensembles.
Hebb also suggested a mechanism responsible for the integration of neu-rons into a resonant structure of an ensemble. “Neurons that fire together, wire together” was a phrase coined by Hebb to explain “associative lear-ning,” in which simultaneous activation of neurons leads to pronounced increases in synaptic strength between those neurons (Hebb, 1949). Hebb wrote: “...any two cells or systems of cells that are repeatedly active at the same time will tend to become ‘associated,’ so that activity in one facilitates activity in the other. ... When one cell repeatedly assists in firing another, the axon of the first cell develops synaptic knobs (or enlarges them if they already exist) in contact with the soma of the second cell.”
(Work by Eric Kandel and others has further provided direct support for the involvement of Hebbian learning in the mechanisms of memory; for review, see Squire & Kandel, 2008).
The neurons within the neuronal ensemble that encode the baseball in the Kreiman experiment have, over time, increased the connection strength between all the cells in the ensemble.In other words,the memory of the baseball is physically stored in connections between the various neurons throughout the brain that encode the baseball. In summary, the neuronal ensemble is a group of neurons with enhanced connections that make them prone to synchronize into a common resonant mode. Once the neurons within a neuronal ensemble self-organize into a synchronous activity, the object encoded by the ensemble is perceived by the person.
This tendency of neuronal ensembles to organize into common resonant activity is essentially responsible for our perception of “meaningful, uni-fied and stable objects” and can be demonstrated by electrical stimulation of the selective neurons at the top of the ‘neuronal ensemble pyramid.’3
Neurosurgeon Wilder Penfield invented a procedure in which it was pos-sible to treat patients with severe epilepsy by destroying the nerve cells in the brain where the seizures originated. Before operating, Penfield stimu-lated the brain with electrical probes while the patients were conscious on the operating table, and observed their responses. In this way, he could more accurately target the responsible areas of the brain, reducing the side-effects of the surgery.Penfield reported that stimulation of the tempo- ral lobes could lead to vivid recall of events from memory (Penfield 1975). 4
Conversely,when non-selective neurons in the primary visual area (area V1, at the bottom of the pyramid) were electrically stimulated,subjects only re- ported seeing a flash of light (Penfield,1975;Kastner,1998).Since neurons in V1 are part of a great many neuronal ensembles,stimulation of one cell does not evoke activation of any complete neuronal ensemble and the subjects do not experience seeing any specific objects or events. A signi-ficantly larger proportion of V1 neurons need to be stimulated to trigger activity of the complete neuronal ensemble encoding an object (as happens when a visually presented object is recognized).
3 These selective neurons in the temporal lobe seem to play an especially important role in bon-ding neurons of the neuronal ensembles into a resonant system. For example, damage (particu-larly bilateral damage) to the fusiform gyrus of the inferior temporal lobe may result in prosopag-nosia (Damasio AR, 1982), a complete inability of a subject to identify faces (prosopagnosia comes from the Greek prosopon meaning “face” and agnosia meaning “non-knowledge”). In the absence of bonding neurons in the fusiform gyrus, the neuronal ensembles encoding faces can-not activate into synchronous firing that is necessary for face recognition. Prosopagnosia is one type of visual agnosia. Patients with visual agnosia are able to attend to the object and perceive the fragment, however they are unable to group the fragments together in order to experience the complete object; they cannot make sense of the object or name it. Visual agnosia was made famous by Oliver Sacks’ essay, The Man Who Mistook His Wife for a Hat (Sacks, 1985). It tells the story of the patient named Dr. P who cannot recognize the common objects around him, including a shoe, a foot, his wife, or even his own face. Because of this complete lack of recognition, Dr. P thinks that his wife’s head is a hat and grabs it in order to put it on his head.
4 See Appendix 3 for a detailed description and a discussion of Penfield’s
observations.
Chapter 1: Object encoding in the brain
15 Part 1. Neuroscience of imagination
1.3 Visual recognition and memory recall of an object activate the same neuronal ensemble
There are two ways in which a neuronal ensemble is normally activated leading to the perception of the particular object encoded by that ensemble:
(1) An image of an object presented to our eyes triggers activation of a neuronal ensemble from the bottom up by stimulating neurons in the pri-mary visual area (V1). This process is referred to as “visual recognition.”
(2) When an object is recalled from memory (with eyes shut) the same neuronal ensemble is activated from the top down. This process is referred to as “memory recall.”

Figure 1.5 Visual recognition and memory recall. Recognition and recall can be defined as: self-organization of a neuronal ensemble associated with an object into a synchronously firing unit. When the process of self-organization is triggered from the base of the pyramid by a few visual cues, a sound,or a familiar smell,it is called “recognition” (bottom-up recognition initiated by firing of neurons at the bottom, or base, of the pyramid). When the self-organization is triggered from the top of the pyramid by associative thinking or active planning, the process is called “visual recall” (top-down recall initiated by the firing of neurons at the top of the pyramid, which activates a mental image of the object).
Kreiman’s experiment recorded from single neurons located at the top of the pyramid, in the medial temporal lobe. These neurons were activated during both the visual recognition of a baseball (left panel) and during its recall from memory (mental image, right panel). There is growing scienti-fic evidence that visual recognition and memory recall share underlying neural processes (Kosslyn, 2006). In other words, it’s not just the neurons in the temporal lobe but rather the complete neuronal ensemble which is activated during visual recognition of a particular object, and which is again re-activated during recall of that object from memory.
Chapter 1: Object encoding in the brain 17
Of course this model is a bit of an oversimplification, but it’s a good starting point, and it holds true at least in some experimental paradigms (as discussed below). In Kreiman’s experiment, this means that when a patient saw the picture of a baseball, the baseball’s neuronal ensemble activated to fire synchronously yielding a perception of the baseball; when the subject’s eyes were closed and the image of the baseball was recalled, it was recalled by activating the same neuronal ensemble to fire synchronously, which again yielded a perception of the baseball.
As mentioned above, Kreiman’s experiment is part of a growing body of scientific experimental evidence that supports the notion that visual recognition and memory recall share underlying neural processes: object perception is mediated by activation of the same neuronal ensemble en-coding the object,independently of whether the physical or recalled object is perceived.Let’s take a quick look at some other notable experimental evidence that supports this seminal notion: Researchers led by Hagar Gelbard-Sagiv and Itzhak Fried recorded from hundreds of neurons in and around the hippocampus of 13 epilepsy patients (Gelbard-Sagiv, 2008). As in the Kreiman experiment, the patients were undergoing ope-rations to treat epilepsy. In the course of the operation, surgeons had to introduce electrodes into the brain in order to locate the source of the sei-zures. During the experiment, the patients were shown a number of short (10 second) television clips consisting of audiovisual sequences. The first time the patients saw the clips, some of their neurons were selectively activated only by certain episodes and not by others. For example, one patient’s neurons were activated by a clip from The Simpsons but not by any other clip including a clip of Michael Jordan playing basketball. As discussed in the previous section, this means that the researchers were able to find just one of the neurons which is part of the neuronal en-semble that encodes The Simpsons. Since the neuronal ensemble representing The Simpsons is different from the one representing Michael Jordan, the scientists were able to record from the single neurons that were exclusively part of one of the neuronal ensemble pyramids and not the other.
... "
Näin "meillä":
http://keskustelu.skepsis.fi/Message/FlatMessageIndex/403774?page=1#405104
Tampereen valeyliopiston haistapaskantiedehöntät eivät halua edes kuulla todellisesta ihmistymisestä mitään
https://www.is.fi/tiede/art-2000007931688.html
Suomalaistutkijat selvittivät: Homo sapiens peittosi neandertalilaiset luovuusgeenien avulla
Tutkijan mukaan luovuusgeenit ovat auttaneet ihmisiä selviämään koronaviruksen kaltaisista vitsauksista.

Terho "Hörhö" Lehtimäki johti tutkimusta Liisa Keltingans-Järvisen kanssa.Kuva: Timo Marttila
Mikko Marttinen
5:41 | Päivitetty 5:42
Homo sapiens on paitsi viisas ihminen, myös luova ihminen.
Tämä selviää Tampereen yliopiston tutkijoiden uudesta tutkimuksesta, jonka tulokset on julkaistu arvostetussa Nature-sarjan Molecular Psychiatry -lehdessä.
Geenitutkimus perustuu suomalaisen Lasten ja nuorten sepelvaltimotaudin riskiteki-jät -tutkimuksen (LASERI) ainutlaatuiseen aineistoon, jota verrattiin simpanssien ja neandertalinihmisten DNA:han.
Tutkijat tunnistivat aineistosta 267 tunnepohjaiseen reagoivuuteen, itsehillintään ja itsetietoisuuteen vaikuttavaa geeniä, jotka löytyvät vain nykyihmisistä.
– Nämä geenit todistavat,että luovuuden kehittyminen oli Homo sapiensin evoluution ja elossa säilymisen salainen ase, jota serkuillamme neandertalinihmisillä ei ollut, professori Terho Lehtimäki sanoo Tampereen yliopiston tiedotteessa.
– Tämä huomionarvoinen geenilöytö avaa ensimmäistä kertaa ikkunan menneisyy-teemme ja valaisee käsitystämme itsetietoisuutemme kehityksen tärkeästä roolista ihmisen evoluutiossa.
Lehtimäen mukaan ainutlaatuiset geenit säätelevät prosesseja, joiden ansiosta nykyihmiset ovat sosiaalisia, luovia kertovassa taiteessa ja tieteessä sekä sukulaislajejaan vastustuskyisempiä ikääntymistä, vammoja ja sairauksia vastaan.
Sen jälkeen, kun Homo sapiensit muuttivat Afrikasta noin 60000 vuotta sitten, he syrjäyttivät vähitellen neandertalinihmiset ja muut ihmislajit. Ne kaikki kuolivat sukupuuttoon noin 40000 vuotta sitten.
– Ihmisen luovuus, sosiaalisuus ja terve pitkäikäisyys syntyivät vastauksena tarpee-seen sopeutua ankariin ja monimuotoisiin olosuhteisiin ja kommunikoida suurissa sosiaalisissa ryhmissä 400000 – 100000 vuotta sitten, Lehtimäki toteaa.
– Tälläkin hetkellä kohtaamme covid-19-pandemian vuoksi samanlaisia haasteita, joihin meidän on sopeuduttava luovasti, kuten esi-isämme oman aikansa haasteisiin.
"Aiheesta" aikaisemmin:
http://keskustelu.skepsis.fi/Message/FlatMessageIndex/401874?page=1#401874
Risto Koivula
15.09.2019 22:03:41
401874 ... "
https://www.kemiamedia.fi/luovuus-auttoi-homo-sapiensia-jaamaan-eloon/
Uutiset | 23.4.2021 | Kemiamedian toimitus
Luovuus auttoi Homo sapiensia jäämään eloon
Siinä missä neandertalilainen ja muut ihmislajit kuolivat noin 40000 vuotta sitten su-kupuuttoon, Homo sapiens säilyi hengissä luovuutensa ja seurallisuutensa ansiosta.
Tähän viittaa Tampereen yliopistossa tehty geenitutkimus.
Tutkijat selvittivät geneettisten verkkojen evoluutiota simpanssista nykyihmiseen ja tunnistivat luovuuden geenit, jotka erottavat meidät muista lajeista.
Geenitutkimus perustuu suomalaisen Laseri-hankkeen (Lasten ja nuorten sepelvaltimotaudin riskitekijät) ainutlaatuiseen aineistoon.
Tutkijat tunnistivat aineistosta 267 tunnepohjaiseen reagoivuuteen, itsehillintään ja itsetietoisuuteen vaikuttavaa geeniä, jotka löytyvät vain nykyihmisistä.
”Nämä geenit todistavat, että luovuuden kehittyminen oli Homo sapiensin evoluution ja elossa säilymisen ’salainen ase’, jota serkuillamme neandertalinihmisillä ei ollut”, sanoo kliinisen kemian professori Terho Lehtimäki yliopiston tiedotteessa. Lehtimäen ja psykologian professorin Liisa Keltikangas-Järvisen lisäksi projektissa oli mukana tutkijoita muualta Suomesta, Yhdysvalloista ja Espanjasta.
Tulokset julkaisi Nature-sarjan Molecular Psychiatry -lehti.
Löydetyt geenit säätelevät ihmisen sopeutumista
Tutkimuksessa tunnistetut 267 geeniä löydettiin suuremmasta 972 geenin ryhmästä, jonka sama ryhmä oli tunnistanut jo aiemmin.
”Nämä 267 geeniä yhdessä vastaavat lähes kaikista periytyvistä temperamentin ja luonteenpiirteiden vaihteluista, jotka säätelevät ihmisen sopeutumista ympäristössä tapahtuviin muutoksiin”, Lehtimäki kertoo.
Arvaamattomia muutoksia ja uhkia ovat esimerkiksi sosiaaliset konfliktit, tarttuvat taudit, ilmaston muuttuminen ja selviytymiseen tarvittavien resurssien saantimahdollisuuksien heikkeneminen.
Ilmentymisen säätely ratkaisee
Geenit, jotka erottavat nykyihmisen neandertalilaisista ja simpansseista, ovat lähes kaikki RNA:sta valmistettuja säätelygeenejä eivätkä DNA:sta valmistettuja proteiineja koodaavia geenejä.
Sapiensille ainutlaatuisista geeneistä 95 prosenttia ei ole proteiinia koodaavia, vaan niihin liittyy enimmäkseen pitkiä koodaamattomia rna:ita geeniverkossa, joka säätelee ihmisen itsetietoisuutta.
”Homo sapiensin, neandertalinihmisten ja simpanssien proteiineja koodaavat geenit ovat lähes kaikki samanlaisia. Se, mikä siis erottaa nämä lajit toisistaan on niiden proteiineja koodaavien geenien ilmentymisen säätely.”
(Kuva Adobe Stock) Taiteilijan näkemys neanderintalinihmisistä. Laji kuoli sukupuuttoon, sillä siltä puuttui nykyihmisen luovuus. "
Molecular Psychiatry (2022) 27:354–376
https://doi.org/10.1038/s41380-021-01097-y
E X P E R T R E V I E W
file:///C:/Users/Huawei/Downloads/s41380-021-01097-y.pdf
Evolution of genetic networks for human creativity
I. Zwir 1, 2
R. Romero-Zaliz 2
A. Mesa 2
J. Arnedo 2
R. Salas 5
G. F. Poblete 5, 6
E. Raitoharju 7
O. Raitakari 8
L. Keltikangas-Järvinen 9
I. Tattersall 11
C. R. Cloninger 1, 4
Received: 30 October 2020 / Revised: 19 March 2021 / Accepted: 31 March 2021 / Published online: 21 April 2021
© The Author(s) 2021. This article is published with open access
C. R. Cloninger
1 Department of Psychiatry, Washington University School of Medicine, St. Louis, MO, USA
2 Department of Computer Science and Artificial Intelligence, University of Granada, Andalusian Research Institute in Data
Science and Computational Intelligence, Granada, Spain
3 Unit of Psychology, Faculty of Education, University of Oulu, Oulu, Finland
4 Anthropedia Foundation, St. Louis, MO, USA
5 The Menninger Clinic,Baylor College of Medicine,and DeBakey VA Medical Center,Houston, TX, USA
6 The Menninger Clinic, Houston, TX, USA
7 Department of Clinical Chemistry, Fimlab Laboratories, and Finnish Cardiovascular Re-search Center - Tampere, Faculty of Medicine and Health Technology, Tampere University, Tampere, Finland
8 Center for Population Health Research, University of Turku and Turku University Hospital; Research Center of Applied and Preventive Cardiovascular Medicine, University of Turku; Department of Clinical Physiology and Nuclear Medicine, Turku University Hospital, Turku, Finland
9 Department of Psychology and Logopedics, University of Helsinki, Helsinki, Finland
10 Department of Psychiatry, University of Texas San Antonio, Long School of Medicine, The Glenn Briggs Institute of Alzheimer’s and Neurodegenerative Disorders, San Antonio, TX, USA
11 American Museum of Natural History, New York, NY, USA
These authors contributed equally: I. Zwir, C. Del-Val
Supplementary information
The online version contains supplementary material available at https://doi.org/10.1038/s41380-021-01097-y.
Abstract
The genetic basis for the emergence of creativity in modern humans remains a mystery des-pite sequencing the genomes of chimpanzees and Neanderthals,our closest hominid relatives. Data-driven methods allowed us to uncover networks of genes distinguishing the three major systems of modern human personality and adaptability: emotional reactivity, self-control, and self-awareness. Now we have identified which of these genes are present in chimpanzees and Neanderthals. We replicated our findings in separate analyses of three high-coverage genomes of Neanderthals. We found that Neanderthals had nearly the same genes for emotio-nal reactivity as chimpanzees,and they were intermediate between modern humans and chim- panzees in their numbers of genes for both self-control and self-awareness. 95% of the 267 genes we found only in modern humans were not protein-coding, including many long-non-coding RNAs in the self-awareness network. These genes may have arisen by positive selec-tion for the characteristics of human well-being and behavioral modernity, in-cluding creati-vity, prosocial behavior, and healthy longevity. The genes that cluster in asso-ciation with those found only in modern humans are over-expressed in brain regions involved in human self-awareness and creativity, including late-myelinating and phylogenetically recent regions of neocortex for autobiographical memory in frontal, parietal, and temporal regions, as well as related components of cortico-thalamo-ponto-cerebellar-cortical and cortico-striato-corti-cal loops. We conclude that modern humans have more than 200 unique non-protein-coding genes regulating co-expression of many more protein-coding genes in coordinated networks that underlie their capacities for self-awareness, creativity, prosocial behavior, and healthy longevity, which are not found in chimpanzees or Neanderthals.
Introduction
One of the most fundamental questions about human nature is what sparked the explosive emergence of creativity in modern humans before their widespread dispersal from Africa and the subsequent extinction of Neanderthals [1–4].
Major controversies persist about the basis for human creativity in art and science, as well as about the differences in cognition, language, and personality that distinguish modern humans from extinct hominids [5–8]. These controversies occur because the archeological and fossil records are incomplete and subject to ambiguous interpretation [9, 10].
What distinguishes behaviorally modern humans from other hominids?
Anthropologists distinguish behaviorally modern Homo sapiens (Sapiens) from other hominids by virtue of Sapiens’ enhanced cognitive, social, and physical adaptability.
Behaviorally modern Sapiens demonstrate remarkable creativity compared to other homi-nids: that is, they show signs of innovation, flexibility, depth of planning, and related cogni-tive abilities for symbolism and self-awareness that enable spontaneous generation of narra-tive art and language [2, 5,11–13]. Early behaviorally modern Sapiens were also more proso- cial in their behaviors than archaic hominids: they maintained larger social groups, estab-lished reciprocal social networks for remote trade, and regularly cooperated with one another in groups composed partially or completely of non-kin, as well as providing altruistic support and cooperation with non-kin who were raising children or disabled [11, 14, 15].
Behaviorally modern Sapiens are also distinguished by their healthy longevity, as evidenced by their resilience to cold and other climatic extremes [16], lower energy requirements and reduced mortality from injury and disease [17–19], and a prolonged post-reproductive lifespan that facilitates cooperative breeding [11, 20, 21], which have all enhanced health and viability in diverse, harsh, and unpredictable habitats throughout the world.
The lineages of Sapiens and Homo neanderthalensis (Neanderthals) are thought to have diverged from a common ancestor during the Middle Pleistocene before 500 thousand years ago (kya), at a time when the lineage of Sapiens was isolated in Africa and that of Neander-thals was confined to Europe and western Asia [22,23]. Precursor forms to Neanderthals are recognized at least by 430 kya in Europe [24],but the behaviors and genomes of Neanderthals themselves are best known from artifacts and fossils dating from 130 to 40 kya in Eurasia [Supplementary Information]. In contrast, anatomically modern Sapiens emerged in eastern Africa [22] by 200 to 160 kya [2, 25] following a period from 320 to 200 kya marked by un-predictable climactic fluctuations [15, 25, 26] that were superimposed on a long-term pat-tern of progressive aridity [27]. Under these challenging ecological conditions, precursors of behaviorally modern Sapiens began to maintain larger social groups and reciprocal social al-liances with non-kin (e.g., remote trade networks), express themselves symbolically (e.g., art, ornamentation), collect remote resources (e.g., pigments, obsidian, and other special stones) for later use, flexibly use expanded dietary options (e.g., fishing and collecting shellfish) in times of unpredictable resource availability, and began to accumulate cultural knowledge and standardized technologies that enhanced their adaptability and well-being [15,25,26, 28].
Recent findings, however, suggest that behaviorally modern Sapiens, with a distinctively more imaginative and flexible set of abilities that had not been observed in any hominids there or elsewhere, emerged in Africa about 100 kya and spread throughout the continent thereafter [29].
The ecological and economic pressure on the smaller bands of mobile and muscular hunters in the lineage of the Neanderthals in Europe were different from those on the lineage of Sapiens in East Africa [30]. Neanderthals and their European ancestors were less resilient to climatic extremes, particularly cold, and their hunting of large land animals demanded high daily energy expenditures [31]. Nevertheless,Neanderthals were able to function successfully before they had to compete with Sapiens.Neanderthals were able to conserve their reliance on hunting large land animals by moving out of inhospitable areas without the need to develop greater social connectedness or more efficient and diverse technologies and resources [17, 22, 28, 31].
The innovative practices of Sapiens are best documented after 50 kya when they flourished to a stage in which the creative imagination of fully modern humans was unmistakably dis-played [2, 5, 32]. The flourishing of behavioral modernity in the late Pleistocene is likely to have been facilitated by incremental cultural and neurobiological processes by which comp-lex behaviors like narrative figural art and language emerged by exaptation and behavioral recruitment [2]. In any case, the basic features of behavioral modernity must have been alrea-dy present when Sapiens spread out of Africa between 65 and 55 kya, while the African cli-mate became drier and colder [27,33]. The inventive, sociable,and resilient Sapiens were able to adapt well to unpredictable and diverse conditions as they migrated out of Africa and spread throughout the world, replacing all other hominids by 40 kya and producing cultures that flourished by continuing to expand in knowledge, art, science, technology, and population density to the present day.
What is creativity? How is it measured?
The most distinctive and prominent feature of behavioral modernity identified by paleoanth- ropologists and archeologists is what psychologists have described as creativity, particularly the achievements and personality traits of Evolution of genetic networks for human creativity highly creative people.
355
Creativity can be succinctly defined as the use of imagination or original ideas to achieve valued goals [34, 35], and is a multifaceted phenomenon that can be assessed in terms of par-ticular aspects of intelligence and/or particular aspects of personality [36-38].The psychomet- ric tests of the creative aspects of intelligence were developed by Guilford and Torrance to measure aspects of divergent thinking in verbal and pictorial tasks. Divergent thinking is an innovative way of solving problems by exploring many possible solutions, making sponta-neous intuitive connections among what are conventionally regarded as disparate phenome-na, while tolerating some ambiguity [39]. Divergent thinking typically occurs in states of restful and playful self-aware evaluation of internal thoughts and images, such as mind-wan-dering in the default mode, flow, free association, day-dreaming, or contemplation [37, 40 – 42], which depends on activation of the medial prefrontal cortex for evaluation of internal stimuli as a core component of the self-awareness network [43, 44]. In contrast, convergent thinking follows a logical sequence of inferences to arrive at a single solution with certainty; it depends on the lateral prefrontal and parietal cortices, which are core components of the executive self-control network that supports purposeful use of symbols and intentional inhibition of externally triggered impulses [19, 45, 46].
The features used to measure divergent thinking include originality (inventive and imagina-tive thoughts), flexibility (ability to move from one conceptual field to another), fluency (free-flow of many relevant ideas and responses), elaboration (many vivid, specific details), a high degree of abstraction, and persistence despite uncertainty [39, 47, 48].
Divergent thinking tests developed by Guilford and Torrance are the most widely used tests of creative intellectual functioning because they are strongly predictive of creative achievement and problem-solving ability in everyday life [38, 47, 48].
How is creativity related to other aspects of behavioral modernity?
In addition to its cognitive properties, divergent thinking involves relaxed states of intuitive awareness that are also characterized by physical spontaneity, cheerful affect, playfulness, and sociability [40, 49, 50], which can be quantified in terms of personality characteristics. Personality refers to the way an individual learns to shape and adapt to an ever-changing internal and external environment [51].
Like divergent thinking, creative personality features are multi-faceted, including character traits (i.e.,styles of rational self-government,with executive functions of self-directedness, legislative functions of cooperativeness, and judicial functions of self-transcendence) and temperament traits (i.e., emotional drives of curiosity about what is novel, willingness to take risks, willingness to work for social recognition, and perseverance for the sake of achieve-ment) [36,52,53].The two domains of temperament and character make it clear that a person’s potential for creativity cannot develop without both the wisdom to recognize what is valuable and the plasticity to adapt accordingly. The most widely used psychometric test for assessing both domains of the creative personality is the Temperament and Character Inventory (TCI) [51, 52, 54].
Tests of creative divergent thinking in verbal and pictorial tasks, creative personality traits as measured by the TCI, and direct assessments of lifetime creative achievements, are each highly reliable and validated by their strong correlations with one another even when general intelligence and demographic variables are controlled [47, 48]. Empirically, the TCI creative personality profile also measures human health in general, including physical, mental, and social well-being [19, 53, 55].
Put another way, the three domains of features of behavioral modernity identified by anthropologists are in fact interdependent aspects of modern human health and well-being.
As a result, the TCI provides valid quantitative phenotypic measures with which to investi-gate the cognitive, emotional, and social functions, brain connectivity, and genetics underly-ing creativity, prosociality, and other aspects of well-being in modern humans in ways that are robustly replicable [19, 56].
Cognitive scientists have proposed that the creative ability of Sapiens to see the world and other people in new ways depends on several interrelated brain processes of learning and memory. Sapiens’ creativity is thought to depend on human brain functions for prospective learning (i.e., the encoding, storing, and retrieval of intended actions), constructive learning (i.e., recollection of the past and imagining the future),and the related capacities for theory of mind (i.e., the ability to attribute mental states to ourselves and others to facilitate empathic social interaction), the default mode (i.e.,awareness of internal milieu without focus on exter- nal tasks), autobiographical memory (i.e., vivid recollection of past experiences with contex-tual awareness of when and where facts were learned),and story-telling (i.e.,meaningful com- position of narrative figural art and language), which are all aspects of self-awareness and share largely overlapping brain circuitry [57–59]. In turn, these processes underlying diver-gent thinking operate cooperatively with other processes underlying convergent thinking, which have complementary functions for problem-solving in successful daily living [39, 46]. Just as creative personality is multifaceted, its related brain functions are also multifaceted aspects of complex neurocognitive systems of adaptability that are measured by the TCI [19].
357
How do complex adaptive abilities develop and evolve?
Complex adaptive traits become organized by developmental [60] and evolutionary [61–63] processes characterized by multi-finality (i.e., the same antecedents can have different out-comes, as in genotypic pleiotropy) and equi-finality (i.e., different antecedents can have the same outcome, as in heterogeneity from redundant genotypic paths), as we have investigated in detail for the genotypic–phenotypic architecture of the TCI [19,64, 65].These properties of complex systems are the basis for the important role of exaptation,that is when already occur- ring characters are co-opted to enable new adaptive functions [66]. Such plasticity permits the creativity to make new things out of old parts, or, more specifically, to produce complex adaptive phenotypes and genotypes via nonlinear dynamical interactions among constituent features to select for advantageous novel functions [23]. The development of such complex adaptive functions are likely to be positively selected in evolution when they are beneficial for survival and reproduction, as has been suggested for the evolution of creativity in Sapiens in response to unpredictable climatic fluctuations and resource variability that threatened survival [26], or when large and cooperative social groups and trade networks began to benefit from enhanced communication by language to facilitate communication [2, 7, 67].
As a result of the incompleteness of the archeological record, there has been substantial cont-roversy about whether the features of behavioral modernity emerged as a full set all at once during the late Pleistocene [2, 5], or if some features emerged individually and/or successive-ly and then became organized in progressive stages in Africa in response to increased envi-ronmental pressure after the common ancestor of Sapiens and Neanderthals had dispersed to Europe before 500 kya [15, 25, 68]. The most recent information suggests the adoption of more complex behaviors after 400 kya and the emergence of the most distinctive features of behavioral modernity after 100 kya [25,26], which suggests the role of many genes in coordi- nated networks, as expected for such complex adaptive traits. The complex behaviors obser-ved before and after 100 kya may be distinguished best by the difference between convergent and divergent thinking because the best documentation of the modern creative imagination is narrative figural art [2, 5, 32], which requires both symbolism and self-awareness. Evidence for such creative divergent thinking first appears after 50 kya, and the creative
achievements of Sapiens continue to accumulate to thisday [11].
In any case, the characteristics of behavioral modernity are certainly complex adaptive traits that cannot be understood by focusing on one brain function or one gene at a time. Unraveling the complexities of behavioral modernity presents many daunting challenges.
Challenges of understanding the evolution of human creativity
Comparisons of Sapiens to other living anthropoid primates provide circumstantial evidence that changes in brain circuitry and related functions for symbolism and/or self-awareness ac-count for the creative characteristics that distinguish behaviorally modern humans from other hominids [12, 45, 57]. However, cranial fossils provide only limited information about the brains of Neanderthals and other extinct hominids [12, 45, 69 – 71]. Archeological evidence indicates that Neanderthal cultures and technologies showed little of the spirit of innovation that animated their counterparts among the Sapiens who replaced them in Europe and wes-tern Asia beginning at some time over 40 thousand years ago [9]. Neanderthals had sophisti-cated executive skills and did produce the occasional expression that might be interpreted as symbolic,but all the artifacts suggesting this is dated after 130 kya [17] and mostly around 40 kya after Sapiens had begun to migrate out of Africa [72-75]. It is clear that the creative use of symbols by Neanderthals was not a routine part of their lives and cultures, and that, al-though undoubtedly complex, the relationship of the Neanderthals to the environment around them — and presumably also to each other — was profoundly different from the one that Sapiens exhibits today [5, 6, 9].
On the other hand, the Neanderthals were very close relatives of Sapiens, and undoubtedly shared some of their behavioral, emotional, and cognitive functions [1-3,12,45].As expressed informally by the paleogeneticist Svante Pääbo, “I want to know what changed in fully mo-dern humans, compared with Neanderthals, that made a difference. What made it possible for us to build up these enormous societies and spread around the globe, and develop the techno- logy that I think no one can doubt is unique to humans? There has to be a genetic basis for that, and it is hiding somewhere in these lists [of nucleotide base pairs of human genomes] [76].”
Pääbo acknowledges that progress in answering this question has been limited for two major reasons: first, a large number of changes in the human genome after its divergence from the common ancestor of humans and chimpanzees 7–10 million years ago (mya), and second, the lack of knowledge of the functional consequences of these changes [1]. Progress has also been limited by a lack of knowledge of the complex genotypic-phenotypic architecture of traits related to human creativity and behavioral modernity:the genes that influence complex aspects of human personality, such as creativity, symbolism, prosociality, and language, are likely to involve many genes acting in coordinated networks, rather than independently [77]. In order to circumvent these problems, we began by characterizing the complex genotypic–phenotypic relationships that describe the architecture of modern human personality using the TCI [19, 64, 65].
358
Genotypic–phenotypic relations underlying behavioral modernity
We evaluated modern human personality using the TCI because it provides highly reliable and empirically validated measures of creativity and other aspects of behavioral modernity that are heritable and neurobiologically grounded, including physical, emotional, social, cog-nitive, and spiritual aspects of well-being [19, 42, 47, 56, 78, 79] as well as, or better than, other available tests [54]. The TCI accounts for two domains of personality based on distinct forms of learning and memory: temperament (i.e., the unconscious component of personality — associatively conditioned habits and emotional reactivity) and character (i.e., the self-re-gulatory components of personality — what people make of themselves intentionally and / or creatively) [51].
It was developed as a comprehensive measure of human personality, and captures the cha-racteristics of behavioral modernity, including creativity and prosocial behavior, as reviewed in the preceding section and elsewhere [19, 42, 47, 48, 54, 56, 78, 79]. The TCI indices of creativity and well-being also predict subjective and objective measures of physical health, including healthy longevity [19, 55, 80].
We used data-driven methods to conduct genome-wide association studies of the TCI in three different samples with different environments and cultures (Finns, Germans, and Koreans). In this way, we were able to deconstruct the complex genotypic–phenotypic networks and environmental interactions underlying modern human temperament and character [64, 65]. These methods properly account for the properties of complex adaptive systems, including pleiotropy and genetic heterogeneity [81] (also see “Methods”).
More specifically, we proceeded in steps to characterize phenotypic-genotypic relationships at multiple levels of organization.
First, in an epidemiologically representative sample of 2149 Finns, we identified sets of single-nucleotide polymorphisms (SNPs) that naturally cluster within particular individuals regardless of phenotype.
Second, we uncovered five clusters of people with distinct configurations of the 13 facets of the self-regulatory domain of human personality (i.e., the character dimensions of Self-directedness, Cooperativeness, and Self-Transcendence) regardless of genotype.
359
Third, we found 42 SNP sets that were significantly associated with the character profiles and identified 727 gene loci. We replicated 95% of the 42 SNP sets in a sample of 902 heal-thy Germans and a sample of 1092 Koreans, as well as their association with the character clusters [64].The character-associated genotypic sets were found to modulate specific mole- cular processes in the brain for intentional goal-setting, self-reflection, empathy, episodic learning and memory, and healthy longevity. Remarkably 68% of the 727 genes associated with character were unique to a single character profile [64]. As a result,there were multiple groups of genes that led to each individual character trait. For example,high self-directedness occurred in different individuals by means of distinct molecular processes of the genotypic networks that depended on particular configurations of Self-directedness with other aspects of character traits. That is, the genes for Self-directedness were different in people with the creative character profile (i.e.,all 3 character dimensions are high so valued goals are unsel- fish, prosocial, and altruistic),the organized profile (Self-directedness and Cooperativeness are high but Self-transcendence is low, allowing for both personally and socially responsive action for mutual benefit but not sacrifice for others),or the resourceful profile (only Self-directedness is high, leading to self-centered motives).
Next, we repeated this process with the 12 facets of human emotional drives (i.e., the tempe-rament dimensions of Novelty Seeking, Harm Avoidance, Reward Dependence, and Persis-tence) [65]. We uncovered three clusters of people with distinct temperament profiles re-gardless of genotype. One cluster was specified by low Novelty Seeking, high Reward De-pendence, and high Persistence, which we designated as the reliable temperament set, as dis-cussed in detail elsewhere [56,82]. The other temperament clusters were the antisocial cluster and the emotionally hypersensitive cluster. 51 SNP sets were significantly associated with temperament clusters. The 736 genes that mapped to these SNP sets were enriched in mole-cular pathways activated by associative conditioning in animals, including the ERK, PI3K, and PKC pathways that are crucial for the modulation of synaptic plasticity and long-term learning involved in associative conditioning of emotional reactivity, social attachment, and persistence. We replicated 90% of the 51 SNP sets for temperament clusters in the healthy German and Korean samples.
The genes we uncovered for temperament and characteroverlapped partially, so we evaluated the organization of the temperament and character clusters jointly. We uncovered three phe-notypic networks that accounted for the joint relations of clusters of temperament traits with clusters of character traits. We designated these joint temperament-character clusters as the
(i) emotional-unreliable network (i.e., people who were highly emotionally reactive with little self-control or creativity),
359
(ii) organized-reliable network (i.e., people with strong self-control of emotional conflicts and goals, but little creativity), and
(iii) creative-reliable network (i.e., people who were highly creative, prosocial, and insightful in appraisal of values and theories) [19]. We found that these phenotypic networks were nearly disjoint (i.e., shared few subjects or phenotypic features) (see Supplementary Fig. 1).
Each of the three phenotypic networks was strongly correlated with a different multi-locus genotypic network (see Supplementary Figs. S2 and S3).
The functions of the genes that mapped to the genotypic networks were found to regulate distinct systems of learning and memory underlying personality:
(i) a multi-locus net-work of 249 genes for regulation of emotional reactivity, associative conditioning, and social attachments, which we designated as the “emotional reactivity” network;
(ii) a multi-locus network of 438 genes for regulation of intentional goal-seeking,such as pur- poseful acquisition of food, manufacture of tools, cooperative team-work, logical analysis, and symbolization, which we designated as the “self-control” network; and
(iii) a genotypic network of 574 genes for episodic learning and autobiographic memory of a person’s life as a narrative with past, present, and future within which the person can explore alternative perspectives with intuitive insight and creative imagination, which we designated as the “self-awareness” network.
It is remarkable that 73% of the 972 genes in these three networks are unique to a single net-work. It is rare to find such a strong separation of clusters specified by such complex sets of phenotypic and genotypic variables [19].
The genes we identified for temperament and character accounted for nearly all the heritabi-lity of personality expected from twin studies [64, 65]. The strong relations of the three temperament-character phenotypic networks to three major genotypic networks for human adaptability provided us with valuable tools for evaluating the evolution of human creativity and other aspects of behavioral modernity by comparing the genomes of chimpanzees (Pan troglodytes) and Neanderthals to those of modern humans.
Hypotheses to be tested
We hypothesized that the three nearly disjoint genotypic networks for human adaptability evolved in successive steps during the evolution of modern human personality. To test this hypothesis, we studied the 972 genes identified for personality in Sapiens, many of which were also found in the genomes of Neanderthals and chimpanzees. While only distantly rela-ted,these are the two species closest to modern humans that have well-characterized genomes comparable to the high-coverage genomes of modern humans [1]. We hypothesized that the three networks differ from each other in Sapiens, Neanderthals, and chimpanzees. Specifically, we hypothesized that
(i) chimpanzees would have genes only in the emotional reactivity network,
(ii) both Neanderthals and Sapiens would share many genes for intentional self-control, which was already evident in their common human lineage, and
(iii) genes found only in Sapiens would be most frequent in the network for creative self-awareness, as previously predicted on the basis of coincident changes in brain and behavior during hominoid evolution [12, 45].
Once we identified the genes that were unique to modern humans from these analyses, we evaluated what types of genes distinguished the three networks. We also evaluated alternative transmission patterns and environmental conditions that may account for the sudden emer-gence of creativity in modern humans. Finally, we examined where the genes for learning and personality that are unique to modern humans are expressed in the brain.
Subjects and methods Modern human subjects
Our sample of Sapiens was the Young Finns Study, an epidemiological study of 2149 healthy Finnish subjects who were assessed in 1997, 2001, 2007, and 2012 [83]. All subjects had thorough standardized genotypic, environmental, and phenotypic assessments, including administration of the TCI [64, 65].
Personality indicators of behavioral modernity, creativity, and well-being
The Finnish subjects completed the TCI with 240 self-reported items using a 5-point Likert scale [84]. The internal consistency of scales and their re-test reliability were strong: r > 0.8 for dimensions and >0.65 for individual subscales between follow-ups, including the 15-year follow-up. The averages of the scales and subscales scores across the four assessment occasions were utilized to reduce measurement error.
All subjects completed the TCI to assess four dimensions of temperament (Harm Avoidance, Novelty Seeking, Reward Dependence, and Persistence) and three dimensions of Character (Self-directedness, Cooperativeness, and Self-transcendence). Each of these dimensions has multiple facets (subscales) measuring the expression of that dimension in different situations. Descriptions of high and low scorers on each dimension and its subscales are presented in Supplementary Table S1.
360
Our prior data-driven analyses of the genotypic–phenotypic architecture of the TCI uncovered a naturally occurring hierarchical structure that is important for understanding the complex relations of phenotypic indicators of genotypic pre-dispositions to human behavioral modernity that can be derived from the TCI, such as creativity, well-being, self-awareness, and self-control, as we have described in detail elsewhere [19, 82] and in Supplementary Information. Human personality can be described at three levels of complexity from
(1) individual temperament and character dimensions, each composed of the sum of their subscales, as shown in Supplementary Table S1,
(2) genetically independent multi-trait temperament profiles or multi-trait character profiles [64, 65], and
(3) joint networks of temperament and character profiles that indicate integration of multiple learning processes from each of the three major systems of human learning and adaptability, as described elsewhere [19] and briefly in the introduction.
We used two indices of health derived from the TCI, an index of well-being and an index of resilience from ill-being. We have confirmed the validity of the creative personality profile in which all three TCI character traits (i.e.,Self-directedness,Cooperativeness,and Self-Trans-cendence) are highly developed as an index of well-being in multiple samples of modern humans in different cultures [19,52, 55, 80]. Likewise, ignoring Self-Transcendence, the sum of Self-directedness and Cooperativeness is an indicator of resilience from ill-being in many cultures [19, 52, 55, 80]. In our sample of Finns we confirmed the validity of these two in-dices with independent measures of positive affective balance, perceived social support, phy-sical behaviors (exercise, smoking, diet), and objective laboratory findings for ideal health as recommended by the American Heart Association, as described elsewhere [19] and in Supplementary Table S2.
Gene annotation
The study has been carried out with 972 genes mapped to the three phenotypic networks: Creative-Reliable, Organized-Reliable, and Emotional-Unreliable (Supplementary Fig. S1) [19]. We refer to the corresponding genotypic networks as the Self-awareness, Self-control, and Emotional Reactivity net-works, respectively (Supplementary Table S1, and Figs. S2, S3). The annotations of individual genes were obtained using the perl API of Ensembl [85] versions 87-92 (Supplementary Table S3) and classified according to their biotype distinguishing between protein-coding genes, non-coding RNA genes, and pseudogenes (Supplementary Table S4).
Comparative genomics
Chimpanzee orthologs for the 972 genes related to personality in modern Homo sapiens were obtained by accessing the CHIMP2.1.4 database, which uses the Pan troglodytes model (7/20/16) built from genome (v. 2.1.4) with gene model files (R.89) from Ensembl using the Perl API [86]. The orthologous genes for other primates (Bonobo, Chimpanzee, Gibbon, Gorilla, Human, Macaque, Marmoset, and Orangutan,) were obtained using programmatic access to resources in Ensembl [87].
Neanderthal orthologs of the 972 genes related to personality in Sapiens were identified in annotated data of the Neanderthal Genome Project [88]. The replicability of these findings was then evaluated in separate analyses of the high-coverage genomes of the Altai Neanderthal [88], another Neanderthal from the Vindija cave [89]
(specimen 33.19, http://cdna.eva.mpg.de/neandertal/Vindija),
and a third from the Chagyrskaya cave [90] (http://cdna.eva.mpg.de/neandertal/Chagyr-skaya). These analyses enabled us to identify genes that chimpanzees and/or Neanderthals shared with modern humans from those that were only found in modern humans (Supple-mentary Table S5) and then to compare their characteristics (Supplementary Tables S6 and S7).
General statistical methods
We used the Analysis of Variance (ANOVA) to test the null hypothesis that the three studied networks are similar in terms of the genes that compose them within one species and across species (Sapiens, Chimpanzee, and Neanderthal). To do so, we utilized both the ANOVA for independent and correlated samples,one per network (Self-awareness vs Self-control vs Emo-tional Reactivity) in each of the species. Then we applied post-ANOVA comparisons using Tukey’s range Honestly Significant Differences (HSD) Test to evaluate the specific diffe-rences between pairs of networks (e.g., Creative vs Organized). We used the ANOVA test as implemented in Concepts & Applications of Inferential Statistics, Richard Lowry 1998 – 2021, http://vassarstats.net/anova1u.html, and in the rstatix package in R.
For clarity in reporting results, p values reported in ANOVA and Tukey’s HSD test descrip-tives were rounded up to more conservative significance values 0.0001 and 0.01, respective- ly, which indeed tend to exceed the E-30 and E-10 values, respectively. The ANOVA effect size was calculated as the f value defined by Cohen [91], where he proposed the following interpretation of this value: f = 0.1 is a small effect, f = 0.25 is a medium effect, and f = 0.4 is a large effect. All other parameters used in each measurement of ANOVA were calculated as usual [91, 92], and full summaries of all our ANOVAs are provided in Supplementary Information and Supplementary Tables S8–S12.
Genotypic estimation of the behavioral modernity of Neanderthals
The number of individual genes that Neanderthals shared with modern humans may not be an adequate indicator of their impact on creativity and other aspects of modern 361 human functioning. We, therefore, evaluated the impact of genes on the predisposition to modern human well-being as an indicator of behavioral modernity by estimating their relative roles in specific SNP sets to take into account the interactions among coordinated sets of genes that impact well-being.
In order to extract prototypical samples of humans with distinctive Neanderthal-like features and distinctive Sapiens-like features, we first identified the genes found only in Sapiens and the genes Neanderthals shared with Sapiens, excluding genes present in chimpanzees (Supplementary Table S3). Then we cross-correlated these genes with the original SNP sets in which they had been detected in relation to character [64] and/or temperament [65] (see Supplementary Information). We selected SNP sets found in the genotypic networks for self-awareness, self-control, and emotional reactivity, for which we already had measured the associated levels of functioning in modern humans, including two indices (well-being and resilience from ill-being) [19]. From the measures of well-being that we had for SNP sets that contained one or more of the genes that Neanderthals shared with Sapiens, we then estimated the mean well-being of Neanderthal-like humans by weighting the well-being of people in those individual SNP sets by the proportion of genes present in Neanderthals compared to Sapiens in that SNP set for each of the net-works. Likewise, we estimated the mean well-being of prototypical Sapiens-like humans from the measures of well-being in SNP sets that contained one or more of the genes found only in modern humans. Finally, we compared the levels of weighted well-being in SNP sets from the Neanderthal-like humans to Sapiens-like human prototypes using ANOVA statistics, including post-ANOVA comparisons and effect sizes. Finally, we estimated the relative genotypic modernity of these prototypes for the two species from the ratio of their mean levels of well-being.
Horizontal gene transfer (HGT)
In order to determine if genes mapped to the three phenotypic networks could have been ho-rizontally acquired, we calculated their overlap to the regions of HGT identified by Huang et al. [88] in the human reference genome hg 19 [93].
Derived allele frequency (DAF)
We compared the DAF scores for long-intronic-non-coding (linc) RNA genes (Supplemen-tary Table S4) found in Neanderthals with those found only in Sapiens to test for differential selection (Supplementary Tables S6 and S7). DAF scores [94] were calculated for lincRNA genes, including their exons and promoters, using the AnnLoc tool (http://annolnc.cbi.pku.edu.cn).
DAF scores of 0.1 or less are associated with reduced diversity indicative of purifying selec-tion (i.e.,negative selection against deleterious alleles), whereas DAF scores >0.1 indicate in- creased diversity, as may occur with nonfunctional alleles, positive selection of advantageous alleles, or addition of new advantageous alleles in genes by HGT [95, 96].
Gene co-expression in brain
To evaluate the functions of the genes we found only in Sapiens further, we used Process Genes List to analyze lists of genes that mapped to particular SNP sets with at least one gene found only in Sapiens [97]. This machine learning method uses the Allen Brain Atlas to calculate a normalized average mRNA expression level in each brain region for lists of each gene set. Brain regions in which those genes were significantly co-expressed were identified and displayed in brain images.
Further details about all our methods and statistical analyses are available as Supplementary Information.
Results
Genotypic personality networks distinguish Sapiens from other hominoids
We first tested which of the 972 genes associated with joint temperament-character networks of Sapiens (Supplementary Table S3) were also present in genomes of Neanderthals and/or chimpanzees. We found 509 genes for modern human personality in all three hominoids, 148 in Neanderthals but not in chimpanzees, 48 in chimpanzees and not in Neanderthals, and 267 only in Sapiens (Table 1).
We hypothesized that the genes that mapped to the genotypic networks for emotional reacti-vity, self-control, and self-awareness would be differentially present in the genomes of chim-panzees, Neanderthals, and Sapiens. To test this, we performed ANOVA for the genes in the three networks along with contrasts of the possible pairs of species under assumptions of cor-related samples (as occurs with vertical inheritance from parent to offspring) or of indepen-dent samples (as occurs with HGT from organisms other than parents), as summarized in Table 2 and described in Supplementary Table S8. We found that the three species differ sig-nificantly from one another for each of the three networks whether or not the samples are correlated (239 > F (2[1664,2913]) > 16, p < 0.0001) (Table 2, Fig. 1, Supplementary Table S8). The differences among the species were large for genes related to self-awareness (Cohen’s effect size f = 0.46), intermediate for self-control (f = 0.35), and small for emotional reactivity (f = 0.21) (Supplementary Table S8).
362
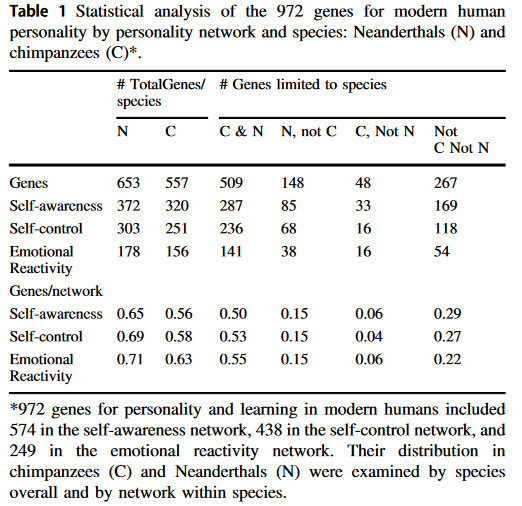
In pair-wise comparisons, Sapiens differed significantly from Neanderthals as well as from chimpanzees in the genes found in each of the three networks, whether or not the samples are correlated (Tukey’s HSD test, p < 0.01) (Table 2, Supplementary Table S8). There was only a small difference between Neanderthals and chimpanzees in the genes they had in each of the three networks; the difference was weakly significant for each of the networks if the samples are considered correlated (Tukey’s HSD test, p < 0.05) and insignificant for the emotional reactivity net-work if the samples are independent (Table 2, Supplementary Table S8). Later we examine the possible occurrence of vertical and horizontal transfer in the evo-lution of human personality, but the differences observed here in Table 2 were about the same whether or not the samples are considered correlated; the one exception was that the smallest difference (i.e., between Neanderthals and chimpanzees in the genes regulating emotional reactivity) was not significant if the samples are considered independent. In sum, we found that the genes present in each of the three genotypic networks differ among each pair of the three hominoid species (Tukey’s HSD test, p < 0.01 to p < 0.05).
Next,we observed that the genes in the three networks are represented in hominoid species as cumulative additions, consistent with the hypothesis that Neanderthals were similar to chim-panzees in their genes for emotional reactivity but were intermediate between chimpanzees and Sapiens in the number of genes present in both the self-control and self-awareness net-works (Fig. 1). Of the 972 genes significantly associated with personality in Sapiens, were present in Neanderthals and 557 in chimpanzees
(Table 1).
653
When compared to chimpanzees Neanderthals did not differ in their proportions of genes for emotional reactivity whether all genes related to personality in each species were considered (Table 1, e.g., 71% of 653, 62% of 557), or the total genes for human personality were accounted (18% vs 16% of 972, F (1, 1972) = 1.76, not significant) (Supplementary Table S9). Putting aside the 54 genes found only in modern humans, 72% of the 195 genes for emotional reactivity were common to all three species.
However, when compared to chimpanzees Neanderthals did have a greater proportion of the genes for self-control (32% vs 25% of 972, F (1, 1942) = 6.86, p < 0.008) and for self-awareness (38% vs 33% of 972, F (1, 1942) = 9.1, p < 0.0001) when all genes for human personality were considered (Supplementary Table S9).
Putting aside the genes for human personality present in chimpanzees, we found Neander-thals had only 33% of the genes in the self-awareness network of Sapiens (viz, 85 of 254 genes), 37% of the genes in self-control network (viz, 68 of 186), 41% of the genes in the emotional reactivity network (viz, 38 of 92). In other words, excluding the genes present in chimpanzees, 67% of genes for self-awareness, 63% of the genes for self-control, and 59% of the genes for emotional reactivity were found only in Sapiens.
Nevertheless, we recognized that the number of genes that Neanderthals shared with Sapiens might not be a direct indicator of their impact on creativity and other aspects of well-being because of the modular organization of genes in complex systems. Specifically, we needed to evaluate the impact of individual genes on well-being by estimating their relative roles in specific SNP sets to take into account the interactions among coordinated sets of genes that impact well-being.
We, therefore, estimated the impact on well-being of the genes shared by Neanderthals and Sapiens in comparison to the impact on well-being of the genes found only in Sapiens. In or-der to extract prototypical samples of humans with distinctive Neanderthal-like features and distinctive Sapiens-like features, we first identified the 267 genes found only in Sapiens and the 148 genes Neanderthals shared with Sapiens,excluding genes present in chimpanzees (Table 1, Supplementary Table S3). We estimated the mean level of well-being of Neander-thal-like humans from the well-being of individuals in naturally occurring clusters of genes (i.e., SNP sets) that included one or more of the 148 distinctive genes of Neanderthals. We estimated the mean level of well-being of prototypical Sapiens-like humans from the well-being of individuals in naturally occurring clusters of genes (i.e., SNP sets) including one or more found only in Sapiens.
363
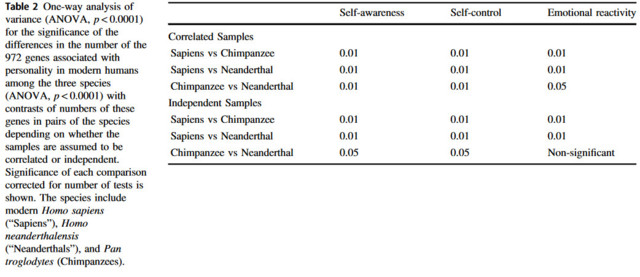
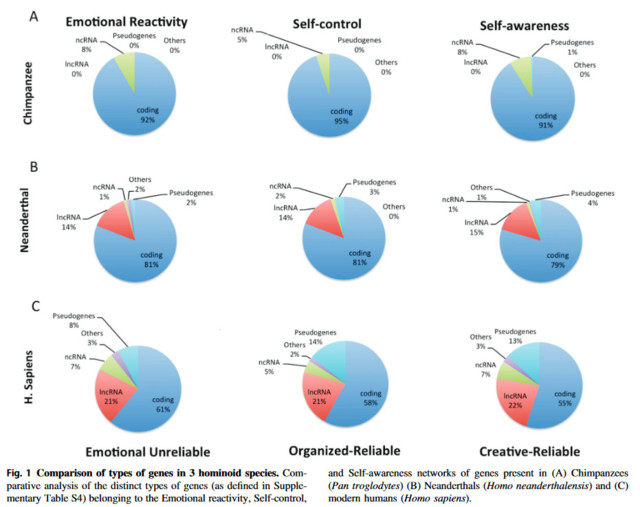
We found that Neanderthal-like groups of genes enhanced well-being more than expected from the number of genes they shared with modern humans, but the mean levels of well-being were still consistently lower for Neanderthal-like humans than for Sapiens-like humans (Supplementary Table S10). The differences between these prototypical groups were similar in comparisons based on two indices of healthy functioning (well-being and resilience from ill-being) for each of the three genotypic networks, including self-awareness and self-control (F (3,252) = 34, F (3,454) = 35, p < 0.0001) and emotional reactivity (F (3, 112) = 15.5, p < 0.0005) (Supplementary Table S10). Specifically, the impact of genes for self-awareness on the well-being of Neanderthal-like humans was 70% of that in Sapiens,which is a rather large difference (effect size f = 0.34). Likewise, the combined impact of genes for self-control and self-awareness on the well-being of Neanderthal-like humans was 67% of that of Sapiens (effect size f = 0.28). The impact of genes for emotional reactivity on the well-being of Neanderthal-like humans was 61% of that of Sapiens (effect size f = 0.20). Similar findings were obtained indicating less resilience to ill-being in Neanderthal-like individuals also (Supplementary Table S10).
Chimpanzees had none of the lncRNAs associated with modern human personality (Fig. 1). Sapiens had more lncRNAs than Neanderthals (21–22% vs 14–15%, Tukey’s HSD test, p < 1E-12) (Fig. 1). The differences in candidate regulatory genes among the hominoid groups were confirmed with the genes present only in Sapiens (Fig. 2).
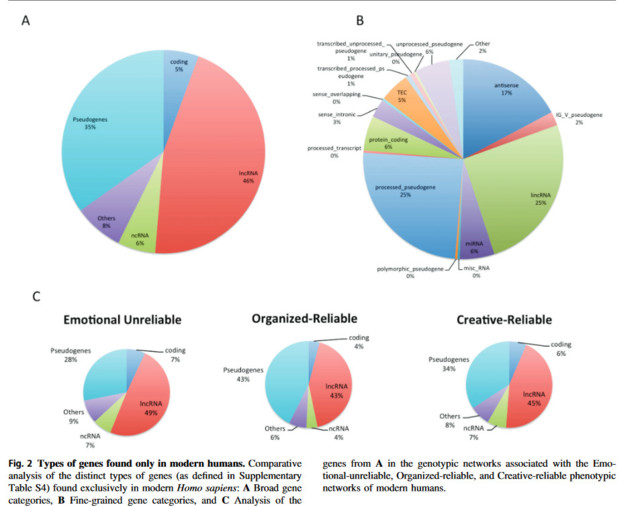
awareness genotypic network (Supplementary Table S5).

MÅÅSES!!! "ROTUMURHAPIERUPAUKKUPEILINEUROONEJA"!!!!
HEWOWWITUN SAATANAN PERKELEEN PASKAA!!!
The mirror neuron system emerges, allowing the understanding of action and the imitation of observed behaviors, a necessary precursor of language.
In the period following 40 thousand years ago the Neanderthals were rapidly replaced in Europe, albeit with some minor gene exchange [117], by invading Homo sapiens whose lives showed unprecedented cultural and technological sophistication. While still itinerant hunter-gatherers, these anatomically and behaviorally distinctive new humans populated the lands-cape in higher densities and brought with them the symbolic tradition of narrative cave art with use of pictorial depth cues in integrated compositions of great complexity and beauty [118]. This innovative practice of creating pictures from the imagination — “the mind’s eye” — is the most powerful indicator we have of the awakening of the modern sensibility, with its profusion of abstract but clearly meaning-laden signs in addition to the sophisticated animal images famous from such localities as Chauvet and Lascaux [73].
The brains of extinct humans are available only as fossil endocasts, limiting the observations that can be made. Compared to chimpanzees, fossil data document the emergence of hemi-spheric asymmetry along with bipedality in australopiths and non-Sapiens. Arising late in hominin history, Neanderthals had large brains that averaged about 1500 ml in volume, more or less identical to those of contemporaneous Pleistocene Homo sapiens (although modern human brains are almost 13% smaller [117]). However, those brains appear to have been or-ganized differently from modern ones: Neanderthals had relatively larger visual areas, while Sapiens have expanded parietal lobes [69] and higher prefrontal regions.On the basis of these findings of coincident differences in brain and behavior,we hypothesized that the genome of Neanderthals would likely be found to have the genetic network for regulation of emotional reactivity and some of the genes of the network for intentional self-control, but not that for self-awareness [12, 45].
Our current findings confirm that the genotypic network for intentional self-control is well-developed in Neanderthals but not in chimpanzees. They also suggest that Neanderthals had acquired genes for self-control and self-awareness in numbers intermediate between mo-dern human and chimpanzees.Excluding genes already present in chimpanzees, Neanderthals had 33% of the 254 genes for self-awareness and 37% of the 186 genes for self-control that are present in Sapiens. Taking into account the modular organization of groups of genes with-in human learning networks, we estimated that the relative level of genotypic predisposition to well-being and modernity of Neanderthal-like humans was 61–70% of that of prototypical Sapiens.
and time. Psychologically,the creative network is so-named because it is found in people who are imaginative, inventive, prosocial, and spiritual [42, 47, 48, 55, 80, 121]. Such self-trans-cendent thinking involves the ability to perceive oneself as a local aspect of a larger spatio-temporal whole, which permits thinking that is free and creative (i.e., “outside the box” of lo-gical deduction and cultural tradition) and theoretically inductive (i.e., extrapolation beyond prior examples based on insight and creative imagination), as expressed in art, science, spiri-tuality, and narrative syntactical language [12, 45]. On the basis of findings of the unique association of coincident changes in brain with cognitive functions for self-awareness and creativity, we hypothesized that only Sapiens were likely to have the genotypic network for self-awareness.
[HM: "Säätiötieteen" tunkiokukkanen...]
2. Tattersall I. Out of Africa: modern human origins special feature: human origins: out of Africa. Proc Natl Acad Sci USA. 2009;106:16018–21.
3. Mellars P. Going east: new genetic and archaeological perspectives on the modern human colonization of Eurasia. Science. 2006; 313: 796–800.
4. Mellars P, Gori KC, Carr M, Soares PA, Richards MB.Genetic and archaeological perspec- tives on the initial modern human colonization of southern Asia. Proc Natl Acad Sci USA. 2013; 110: 10699–704.
5. Klein RG. The human career: human biological and cultural origins. 3rd edn. University of Chicago Press: Chicago, 2009.
6. Tattersall I. An evolutionary framework for the acquisition of symbolic cognition by Homo sapiens. Comp Cognition Behav Rev. 2008; 3: 99–114.
7. Hauser MD, Yang C, Berwick RC, Tattersall I, Ryan MJ, Watumull J, et al. The mystery of language evolution. Front Psychol. 2014;5:401.
8. Elias S. (ed). Origins of human innovation and creativity. Elsevier: Amsterdam, 2012.
9. Tattersall I.Masters of the planet: the search for our human origins. St. Marti’s Press, 2012.
10. Lopez S, van Dorp L, Hellenthal G. Human dispersal out of Africa: a lasting debate. Evol Bioinform Online. 2015;11:57–68.
11. Hill K, Barton M, Hurtado AM. The emergence of human uniqueness: characters underlying behavioral modernity. Evolut Anthropol. 2009;18:187–200.
12. Cloninger CR. The evolution of human brain functions: the functional structure of human consciousness. Aust NZ J Psychiatry. 2009;43:994–1006.
13. Suddendorf T, Addis DR, Corballis MC. Mental time travel and the shaping of the human mind. Philos Trans R Soc Lond B Biol Sci. 2009;364:1317–24.
14. Sussman RW, Cloninger CR. (eds). Origins of cooperation and altruism. Springer: New York, 2011.
15. Brooks AS, Yellen JE, Potts R, Behrensmeyer AK, Deino AL, Leslie DE, et al. Long-distance stone transport and pigment use in the earliest Middle Stone Age. Science. 2018;360:90–94.
16. Staubwasser M, Dragusin V, Onac BP, Assonov S, Ersek V, Hoffmann DL, et al. Impact of climate change on the transition of Neanderthals to modern humans in Europe. Proc Natl Acad Sci USA. 2018;115:9116–21.
17. Anton SC, Potts R, Aiello LC. Evolution of early Homo: an integrated biological perspective. Science. 2014;345:45.
18. Spikins P, Needham A, Wright B, Dytham C, Gatta M, Hitchens G. Living to fight another day: The ecological and evolutionary significance of Neanderthal healthcare. Quarternary Sci Rev. 2019; 217: 98 – 118.
19. Zwir I, Arnedo J, Del-Val C, Pulkki-Råback L, Konte B, Yang SS, et al. Three genetic-environmental networks for human personality. Mol Psychiatry 2019.
20. Lee PC, Fishlock V, Webber CE, Moss CJ. The reproductive advantages of a long life: longevity and senescence in wild female African Elephants. Behav Ecol Sociobiol. 2016; 70: 337 – 45.
21. Croft DP, Brent LJN, Franks DW, Cant MA. The evolution of long life after reproduction. Trends Ecol Evolut. 2015;30:407–16.
22. Hublin JJ. The origin of Neandertals. Proc Natl Acad Sci USA. 2009;106:16022–16027.
23. Stringer C. The origin and evolution of Homo sapiens. Philos Trans R Soc Lond B Biol Sci. 2016; 371:1– 12.
24. Arsuaga JL, Martínez I, Arnold LJ, Aranburu A, Gracia-Téllez A, Sharp WD,et al. Nean- dertal roots:cranial and chronological evidence from Sima de los Huesos. Science. 2014; 344: 1358 – 63.
25. Deino AL, Behrensmeyer AK, Brooks AS, Yellen JE, Sharp WD, Potts R. Chronology of the Acheulean to Middle Stone Age transition in eastern Africa. Science. 2018; 360: 95 – 98.
26. Potts R, Behrensmeyer AK, Faith JT, Tryon CA, Brooks AS,Yellen JE,et al. Environmen- tal dynamics during the onset of the Middle Stone Age in eastern Africa. Science. 2018; 360: 86 – 90.
27. Owen RB, Muiruri VM, Lowenstein TK, Renaut RW, Rabideaux N, Luo S, et al. Progres-sive aridification in East Africa over the last half million years and implications for human evolution. Proc Natl Acad Sci USA. 2018; 115: 11174 – 11179.
28. Marean CW, Bar-Matthews M, Bernatchez J, Fisher E, Goldberg P, Herries AIR, et al. Early human use of marine resources and pigment in South Africa during the Middle Pleistocene. Nature. 2007; 449: 905 – 908.
29. Rito T, Vieira D, Silva M, Conde-Sousa E, Peira L, Mellars P, et al. A dispersal of Homo sapiens from southern to eastern Africa immediately preceded the out-of-Africa migration. NatSci Rep. 2019; 9.
30. Stiner MC, Kuhn SL. Changes in the “connectedness” and resilience of paleolithic societies in Mediterranean ecosystems. Hum Ecol. 2006; 34: 693 – 712.
31. Froehle AW, Churchill SE. Energetic competition between Neandertals and anatomically modern humans. Paleo Anthropology. 2009; 2009: 96 – 116.
32. Brumm A, Oktaviana AA, Burhan B, Hakim B, Lebe R, Zhao J-X, et al. Oldest cave art found in Sulawesi. Sci Adv. 2021;7: eabd4648.
33. Tierney JE, deMenocal PB,Zander PD.A climatic context for the out-of-Africa migration. Geology. 2017; 45: 1023 – 1026.
34. Stevenson A, Lindberg CA (eds). The New Oxford American Dictionary. Oxford University Press: New York, 2010.
35. Sternberg RJ. (ed). The nature of creativity. Cambridge University Press: Cambridge UK, 1988.
36. Sternberg RJ. A three-facet model of creativity. In: Sternberg RJ (ed). The nature of creativity. Cambridge University Press: Cambridge UK, 1988, pp 125–47.
37. Vernon PE (ed). Creativity. Penguin: Baltimore, 1970.
38. Sternberg RJ. The nature of creativity. Creativity Res J. 2006; 18: 87 – 98.
39. Guilford JP. Traits of creativity. In: Anderson HH (ed). Creativity and its cultivation. Harper: New York, 1959, pp 142-61.
40. Freud S. Creative writers and day-dreaming. In: Strachey J (ed). Standard edition of the complete psychological works of Sigmund Freud, vol. 9. Hogarth Press: London, 1908, pp 141 – 54.
41. Rogers CR.Towards a theory of creativity.ETC:A Rev Gen Semant. 1954;11:249–60. 374
43. Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci USA. 2001;98:676–82.
44. Gusnard DA, Akbudak E, Shulman GL, Raichle ME. Medial prefrontal cortex and self-referential mental activity: relation to a default mode of brain function. Proc Natl Acad Sci USA. 2001; 98: 4259 – 64.
45. Cloninger CR, Kedia S. The phylogenesis of human personality: Identifying the pre-cursors of cooperation, altruism, & well-being. In: Sussman RW, Cloninger CR, (eds). The origins of cooperation and altruism. First edn. New York: Springer; 2011. p. 63 – 110.
46. Shen K-K, Welton T, Lyon M, McCorkidale AN, Sutherland GT, Burnham S, et al. Struc-tural core of the executive control network:a high angle resolution diffusion MRI study. Hum Brain Mapp. 2020; 41: 1226–36.
47. Chavez-Eakle RA, Lara MDC, Cruz-Fuentes C. Personality: a possible bridge between creativity and psychopathology? Creativity Res J. 2006; 18: 27 – 38.
48. Takeuchi H, Taki Y, Sekiguchi A, Hashizume H, Nouchi R, Sassa Y,et al.Mean diffusivity of globus pallidus associated with verbal creativity measured by divergent thinking and crea-tivity-related temperaments in young healthy adults. Hum Brain Mapp. 2015; 36: 1808 – 27.
49. Lieberman JN. Playfulness and divergent thinking: an investigation of their relationship at the kindergarten level. J Genet Psychol. 1965; 107: 219 – 24.
50. Vosburg SK. The effects of positive and negative mood on divergent-thinking performance. Creativity Res J. 1998; 11: 165 – 72.
51. Cloninger CR, Svrakic DM, Przybeck TR. A psychobiological model of temperament and character. Arch Gen Psychiatry. 1993; 50: 975 – 90.
52. Cloninger CR. What makes people healthy, happy, and fulfilled in the face of current world challenges? Mens Sana Monogr. 2013; 11: 16 – 24.
53. Cloninger CR, Cloniger KM. People create health:effective health promotion is a creative process. Int J Pers Cent Med. 2013; 3: 114 – 22.
54. Grucza RA, Goldberg LR.The comparative validity of 11 modern personality inventories: predictions of behavioral acts, informant reports, and clinical indicators. J Personal Assess. 2007; 89: 167 – 87.
55. Cloninger CR, Zohar AH. Personality and the perception of health and happiness. J Affect Disord. 2011; 128: 24 – 32.
56. Cloninger CR,Cloninger KM,Zwir I, Keltigangas-Jarvinen L. The complex genetics and biology of human temperament: a review of traditional concepts in relation to new molecular findings. Trans Psychiatry 2019.
57. Tulving E.Episodic memory and common sense: how far apart? Philos Trans R Soc Lond B Biol Sci. 2001; 356: 1505 – 15.
58. Ambrose SH. Coevolution of composite-tool technology, constructive memory, and lan-guage: implications for the evolution of modern human behavior. Curr Anthropol. 2010; 51: 135 – 47.
59. Spreng RN, Mar RA, Kim ASN. The common neural basis of autobiographical memory, prospection, navigation, theory of mind, and the default mode: a quantitative meta-analysis. J Cogn Neurosci. 2008; 21: 489 – 510.
60. Cicchetti D, Rogosch FA. Equifinality and multifinality in developmental psychopatology. Dev Psychopathol. 1996; 8: 597 – 600.
61.Wright S.The shifting balance theory and macroevolution.Annu Rev Genet.1982;16:1–19.
63. Hill WG, Zhang XS. On the pleiotropic structure of the genotype-phenotype map and the evolvability of complex organisms. Genetics. 2012; 190: 1131 – 1137.
64. Zwir I, Arnedo J, Del-Val C, Pulkki-Raback L, Konte B, Yang SS, et al. Uncovering the complex genetics of human character. Mol Psychiatry. 2020; 25: 2295 – 312.
65. Zwir I, Arnedo J, Del-Val C, Pulkki-Raback L, Konte B, Yang SS, et al. Uncovering the complex genetics of human temperament. Mol Psychiatry. 2020; 25: 2275 – 94.
66. Gould SJ, Vrba ES. Exaptation — a missing term in the science of form. Paleobiology. 1982; 8: 4 – 15.
67. Hauser MD, Chomsky N, Fitch WT. The faculty of language: what is it,who has it, and how did it evolve? Science. 2002; 298: 1569 – 79.
68. McBrearty S, Brooks AS. The revolution that wasn’t: a new interpretation of the origin of modern human behavior. J Hum Evol. 2000; 39: 453 – 563.
69. Pearce E, Stringer C, Dunbar RI. New insights into differences in brain organization bet-ween Neanderthals and anatomically modern humans. Proc Biol Sci. 2013; 280: 20130168.
70. Bruner E, Ogihara N, Tanabe HC (eds). Digital endocasts:from skulls to brains. Springer: Tokyo, 2018.
71. Akazawa T, Ogihara N, Tanabe HC, Terashima H (eds).Dynamics of learning in Neander- thals and modern humans: cognitive and physical perspectives. Springer: Tokyo, 2014.
72. Soressi M, McPherron SP, Lenoir M, Dogandzic T, Goldberg P, Jacobs Z, et al. Neander-tals made the first specialized bone tools in Europe. Proc Natl Acad Sci USA. 2013; 110: 14186 – 90.
73. Radovcic D, Srsen AO, Radovcic J, Frayer DW. Evidence for Neandertal jewelry: modified white-tailed eagle claws at Krapina. PLoS One. 2015;10:e0119802.
74. Hoffmann DL, Standish CD, Garcia-Diez M, Pettitt PB, Milton JA, Zilhao J, et al. U-Th dating of carbonate crusts reveals Neandertal origin of Iberian cave art. Science. 2018; 359: 912 – 915.
75. Rodriguez-Vidal J, d’Errico F, Giles Pacheco F, Blasco R, Rosell J, Jennings RP, et al. A rock engraving made by Neanderthals in Gibraltar. Proc Natl Acad Sci USA. 2014; 111: 13301 – 13306.
77. DeSalle R, Tattersall I. What aDNA can (and cannot) tell us about the emergence of language and speech. J Lang Evolut. 2018; 3: 59 – 66.
78. Quoidbach J, Hansenne M, Mottet C. Personality and mental time travel: a differential approach to autonoetic consciousness. Conscious Cognition. 2008; 17: 1082 – 92.
79. Van Schuerbeek P, Baeken C, De Raedt R, De Mey J, Luypaert R. Individual differences in local gray and white matter volumes reflect differences in temperament and character: a voxel-based morphometry study in healthy young females. Brain Res. 2011; 1371: 32 – 42.
80. Zohar AH, Cloninger CR, McCraty R. Personality and heart rate variability: exploring pathways from personality to cardiac coherence and health. J Soc Sci. 2013;1:32–39.
81. Zwir I, Mishra P, Del-Val C, Gu CC, de Erausquin G, Lehtimäki T, et al. Uncovering the complex genetics of human personality: response from authors on the PGMRA model. Mol Psychiatry. 2019.
82. Moreira PAS, Inman RA, Cloninger CR. Personality networks and behavioral problems: Integrating temperament and character using latent profile and latent class analysis. Child Psychiatry Hum Dev. 2020; 2020: 1 – 13.
84. Ravaja N, Keltikangas-Jarvinen K. Cloninger’s temperament and character dimensions in young adulthood and their relation to characteristics of parental alcohol use and smoking. J Stud Alcohol. 2001; 62: 98 – 104.
85. Yates A, Akanni W, Amode MR, Barrell D, Billis K, Carvalho-Silva D, et al. Ensembl 2016. Nucleic Acids Res. 2016; 44: D710 – 716.
86. Smedley D, Haider S, Durinck S, Pandini L, Provero P, Allen J, et al. The BioMart com-munity portal: an innovative alternative to large, centralized data repositories. Nucleic Acids Res. 2015; 43: W589 – 598.
87. Aken BL, Achuthan P, Akanni W, Amode MR, Bernsdorff F, Bhai J, et al. Ensembl 2017. Nucleic Acids Res. 2017;45: D635 – D642.
88. Huang W, Tsai L, Li Y, Hua N, Sun C, Wei C. Widespread of horizontal gene transfer in the human genome. BMC Genom. 2017;18:274.
89. K Prüfer C Filippo SG Mafessoni F Korlević PM Hajdinjak et al. 2017 A high-coverage Neandertal genome from Vindija Cave in Croatia Science 358 655 658.
90. Mafessoni F, Grote S, de Filippo C, Slon V, Kolobova KA, Viola B,et al.A high-coverage Neandertal genome from Chagyrskaya Cave. Proc Natl Acad Sci USA. 2020; 117: 15132 – 15136.
91. Cohen J. Statistical power analysis for the behavioral sciences. 2nd edn. Lawrence Erlbaum: Hillsdale, NJ, 1988.
92. Zhang Z, Yuan K-H. Practical statistical power analysis using Webpower and R. ISDSA Press: Granger, IN, 2018, pp. 356 .
93. Lander ES, Linton LM, Birren B, Nusbaum C, Zody MC, Baldwin J, et al. Initial sequencing and analysis of the human genome. Nature. 2001;409:860–921.
94. 1000 Genomes Project C,Abecasis GR, Altshuler D,Auton A, Brooks LD, Durbin RM, et al. A map of human genome variation from population-scale sequencing. Nature. 2010; 467: 1061 – 73.
95. Ward LD, Kellis M. Evidence of abundant purifying selection in humans for recently acquired regulatory functions. Science. 2012;337:1675–1678.
96. Biswas S, Akey JM. Genomic insights into positive selection.Trends Genet.2006; 22: 437 – 46.
97. Poblete GF, Gosnell SN, Meyer M, Fang M, Nyugen T, Patriquin MA, et al.Process genes list: an approach to link genetics and human brain imaging. J Neurosci Methods. 2020; 339.
98. Mineka S, Suomi SJ. Social separation in monkeys. Psychol Bull. 1978;85:1376–1400.
99. Bowlby J. Attachment. Second edn. Basic Books: New York, 1983.
100. de Waal FB. Primates – a natural heritage of conflict resolution. Science. 2000; 289: 586 – 90.
101. de Waal FB. Cultural primatology comes of age. Nature. 1999; 399: 635 – 636.
102. Whiten A, Spiteri A, Horner V, Bonnie KE, Lambeth SP,Schapiro SJ,et al. Transmission of multiple traditions within and between chimpanzee groups. Curr Biol. 2007;17: 1038 – 43.
103. Sussman RW,Chapman AR (eds). The origins and nature of sociality. Aldine de Gruyter: New York, 2004.
104. de Waal FB. Macaque social culture: development and perpetuation of affiliative networks. J Comp Psychol. 1996; 110: 14 7– 54.
105. Suddendorf T, Whiten A. Mental evolution and development: evidence for secondary representation in children, great apes, and other animals. Psychol Bull. 2001; 127: 629 – 50.
106. Donald M. Origins of the modern mind: three stages in the evolution of culture and cognition. Harvard University Press: Cambridge, MA, 1991.
108. Povinelli DJ, Giambrone S.Reasoning about beliefs: a human specialization? Child Dev. 2001; 72: 691 – 695.
109. McPherron SP, Alemseged Z, Marean CW, Wynn JG, Reed D,Geraads D,et al. Evidence for stone-tool-assisted consumption of animal tissues before 3.39 million years ago at Dikika, Ethiopia. Nature. 2010; 466: 857 – 60.
110. Castellano S, Parra G, Sanchez-Quinto FA, Racimo F, Kuhlwilm M, Kircher M, et al. Patterns of coding variation in the complete exomes of three Neandertals. Proc Natl Acad Sci USA. 2014; 111: 6666 – 71.
111. Niven L, Steele TE, Rendu W, Mallye JB, McPherron SP, Soressi M, et al. Neandertal mobility and large-game hunting: the exploitation of reindeer during the Quina Mousterian at Chez-Pinaud Jonzac (Charente-Maritime, France). J Hum Evol. 2012; 63: 624 – 35.
112. Sandgathe DM, Dibble HL,Goldberg P,McPherron SP, Turq A, Niven L, et al. Timing of the appearance of habitual fire use. Proc Natl Acad Sci USA. 2011;108:E298. author reply E299
113.Tattersall I.The world from beginnings to 4000 BCE.Oxford University Press:New York, 2008b.
114. Sandgathe DM, Dibble HL, Goldberg P, McPherron SP. The Roc de Marsal Neandertal child: a reassessment of its status as a deliberate burial. J Hum Evol. 2011; 61: 243 – 53.
115. Hagen MA. Development of ability to perceive and produce pictorial depth cue of overlapping. Percept Mot Skills. 1976; 42: 1007 – 14.
116. Prufer K, Racimo F, Patterson N, Jay F, Sankararaman S, Sawyer S, et al. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature. 2014; 505: 43 – 49.
117. Green RE, Krause J,Briggs AW, Maricic T,Stenzel U, Kircher M, et al. A draft sequence of the Neandertal genome. Science. 2010; 328: 710 – 22.
118. Brooks KR.Depth perception and the history of three-dimensional art: who produced the first stereoscopic images? I perception. 2017; 8: 2041669516680114.
119. Fu Q, Li H, Moorjani P, Jay F, Slepchenko SM, Bondarev AA, et al. Genome sequence of a 45,000-year-old modern human from western Siberia. Nature. 2014; 514: 445 – 449.
120. Levine B. Autobiographical memory and the self in time: brain lesion effects, functional neuroanatomy, and lifespan development. Brain Cognition. 2004; 55: 54 – 68.
121. Moll J, Krueger F, Zahn R, Pardini M, de Oliveira-Souza R, Grafman J. Human fronto-mesolimbic networks guide decisions about charitable donation. Proc Natl Acad Sci USA. 2006; 103: 15623 – 15628.
122. Ulitsky I. Evolution to the rescue: using comparative genomics to understand long non-coding RNAs. Nat Rev Genet. 2016; 17: 601 – 614.
123. Fang Y, Fullwood MJ. Roles, functions, and mechanisms of long non-coding RNAs in cancer. Genomics Proteom Bioinforma. 2016;14:42–54.
124. Quinn JJ, Chang HY. Unique features of long non-coding RNA biogenesis and function. Nat Rev Genet. 2016; 17: 47 – 62.
125. Lea A, Subramaniam M, Ko A, Lehtimäki T, Raitoharju E, Kahonen M, et al. Genetic and environmental perturbations lead to regulatory decoherence. eLife. 2019; 8: e40538.
126. Rilling JK,Goldsmith DR, Glenn AL,Jairam MR, Elfenbein HA, Dagenais JE, et al. The neural correlates of the affective response to unreciprocated cooperation. Neuropsychologia. 2008; 46: 1256 – 1266.
127. Coxworth JE, Kim PS, McQueen JS, Hawkes K. Grand-mothering life histories and human pair bonding. Proc Natl Acad Sci USA. 2015; 112: 11806 – 11811.
129. Kim PS, Coxworth JE, Hawkes K. Increased longevity evolves from grandmothering. Proc Biol Sci. 2012; 279: 4880 – 4884.
130. Hawkes K, Kim PS, Kennedy B, Bohlender R, Hawks J. A reappraisal of grandmothe-ring and natural selection. Proc Biol Sci. 2011; 278: 1936 – 1938. discussion 1939-1941
131. Schwarz F, Springer SA, Altheide TK,Varki NM, Gagneux P,Varki A. Human-specific derived alleles of CD33 and other genes protect against postreproductive cognitive decline. Proc Natl Acad Sci USA. 2016; 113: 74 – 79.
132. Tattersall I. Brain size and the emergence of modern human cognition In: Schwartz JH (ed). Rethinking human evolution, First edn. MIT Press:Cambridge, MA, 2018, pp 319 - 334.
133. Scally A. The mutation rate in human evolution and demographic inference. Curr Opin Genet Dev. 2016; 41: 36 – 43.
134. Kaessmann H. Origins, evolution, and phenotypic impact of new genes. Genome Res. 2010; 20: 1313 – 1326.
135. King MC, Wilson AC. Evolution at two levels in humans and chimpanzees. Science. 1975; 188:107 – 116.
136. Carroll SB. Evolution at two levels: on genes and form. PLoS Biol. 2005; 3: e245.
137. Gilad Y, Oshlack A, Rifkin SA. Natural selection on gene expression. Trends Genet. 2006; 22: 456 – 461.
139. Crisp A, Boschetti C, Perry M, Tunnacliffe A, Micklem G. Expression of multiple hori-zontally acquired genes is a hallmark of both vertebrate and invertebrate genomes. Genome Biol. 2015; 16: 50.
140. Schrider DR, Navarro FC, Galante PA, Parmigiani RB, Camargo AA, Hahn MW, et al. Gene copy-number polymorphism caused by retrotransposition in humans. PLoS Genet. 2013; 9: e1003242.
141. Navarro FC, Galante PA. A genome-wide landscape of retrocopies in primate genomes. Genome Biol Evol. 2015; 7: 2265 – 2275.
142. Barbash S, Simchovitz A, Buchman AS, Bennett DA, Shifman S, Soreq H. Neuronal-expressed microRNA-targeted pseudo-genes compete with coding genes in the human brain. Transl Psychiatry. 2017; 7: e1199.
143. Kaneko T, Tomonaga M.The perception of self-agency in chimpanzees (Pan trogloytes). Proc R Soc B. 2011; 278: 3694 – 3702.
144. Povinelli DJ. Can comparative psychology crack its toughest nut? Anim Behav Cognition. 2020; 7: 589 – 652.
145. Zohar AH, Zwir I, Wang J,Cloninger CR,Anokhin AP.The development of temperament and character during adolescence: the processes and phases of change. Dev Psychopathol. 2019; 31: 601 – 617.
https://doi.org/10.1038/s41380-019-0399-z
Uncovering the complex genetics of human personality: response from authors on the PGMRA Model
Igor Zwir 1,2
Pashupati Mishra 3
Coral Del-Val 2
C. Charles Gu4
Gabriel A. de Erausquin 5
Terho Lehtimäki 3
C. Robert Cloninger 1,6
Received: 10 February 2019 / Accepted: 14 February 2019
© The Author(s) 2019. This article is published with open access
[email protected]
1 Washington University School of Medicine, Department of Psychiatry, St. Louis, MO, USA
2 University of Granada, Department of Computer Science, Granada, Spain
3 Department of Clinical Chemistry, Fimlab Laboratories, and Finnish Cardiovascular Research Center - Tampere, Faculty of
Medicine and Health Technology, Tampere University, Tampere, Finland
4 Washington University, School of Medicine, Division of Biostatistics, St. Louis, MO, USA
5 University of Texas Rio-Grande Valley, School of Medicine, Department of Psychiatry and Neurology, and Institute of
Neurosciences, Harlingen, TX, USA
6 Washington University, School of Arts and Sciences, Department of Psychological and Brain Sciences, and School of Medicine, Department of Genetics, St. Louis, MO, USA
hypothesis that the heritability expected from twin studies but unexplained by genetic studies is distributed in heterogeneous partitions of a complex trait, each with distinct genotypic-phe-notypic associations. We designed a machine learning algorithm termed PGMRA [1, 3, 4] to identify naturally occurring partitions in the data in an unsupervised fashion. PGMRA first dissects genome-wide data and uncovers a genotypic architecture composed of sets of SNPs shared by subsets of individuals (i.e.,SNP sets [3,6]). Next,phenotypic data are independently organized into natural sets of features such as clinical manifestations [4], voxels of neuroimages [7], or personality traits [1, 2] in a phenomic-like approach [8].
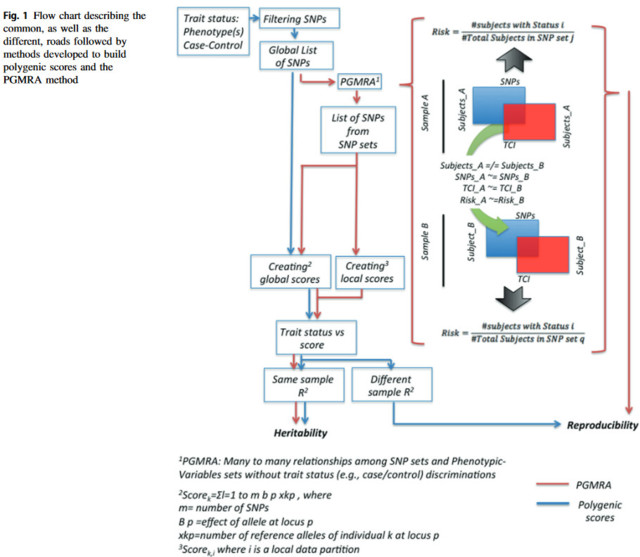
YLE LYKKÄÄ PASKAA KUTEN AINA:
https://yle.fi/a/3-11894011
Suomalaiset tutkijat löysivät nykyihmisen uuden salaisen aseen, jota neandertalinihmisellä ei ollut: 267 ainutlaatuista geeniä
Uusi löytö auttaa tutkijoiden mukaan ymmärtämään, miksi nykyihminen pystyi sopeutumaan ja syrjäyttämään neandertalinihmiset.
Neandertalin ihmisiä kuvaavia malleja saksalaisessa Neanderthal Museum of Mettmannissa vuonna 2009. Kuva: Federico Gambarini / AOP
Miksi nykyihminen ei kuollut sukupuuttoon kuten neandertalinihminen?
Tutkijat uskovat löytäneensä nyt uuden salaisen aseen. Kyseessä on 267 tunnepohjaiseen reagoivuuteen, itsehillintään ja itsetietoisuuteen vaikuttavaa geeniä, jotka löytyvät vain nykyihmisistä.
Geenitutkimus perustuu suomalaisen Lasten ja nuorten sepelvaltimotaudin riskitekijät -tutkimuksen aineistoon.
Tampereen yliopiston tutkijat tunnistivat luovuuden ja selviämisen geenit, jotka erottavat nykyihmiset neandertalinihmisestä ja simpansseista.
– Nämä geenit todistavat, että luovuuden kehittyminen oli Homo sapiensin evoluution ja elossa säilymisen ’salainen ase’, jota serkuillamme neandertalinihmisillä ei ollut, professori Terho Lehtimäki Tampereen yliopistosta sanoo tutkimustiedotteessa.
Yle tavoitti keskiviikkona Lehtimäen Lapin hiihtoladulta. Professori ja Fimlabin ylilääkäri pitää löydöstä merkittävänä, koska pitkään luultiin, ettei asiaa voisi selvittää. Nykytekniikan avulla se oli kuitenkin mahdollista.
Monialainen tutkimus
Professori Terho Lehtimäki ja professori emerita Liisa Keltikangas-Järvinen toteuttivat projektin useiden kollegoidensa kanssa Suomessa, Yhdysvalloissa ja Espanjassa.
Tutkimus yhdistää molekyyligenetiikan, psykologian, neurotieteen, antropologian ja tietojenkäsittelyn uusimmat menetelmät ja käyttää esimerkiksi tekoälyä.
Nyt tunnistetut 267 geeniä löydettiin suuremmasta 972 geenin ryhmästä, jotka sama tutkimusryhmä oli tunnistanut jo aiemmin.
Nämä 267 geeniä vastaavat Lehtimäen mukaan lähes kaikista periytyvistä temperamentin ja luonteenpiirteiden vaihteluista. Tämä taas säätelee esimerkiksi sitä, miten ihminen sopeutuu erilaisiin uhkiin, kuten vaikka koronavirukseen tai ilmastonmuutokseen.
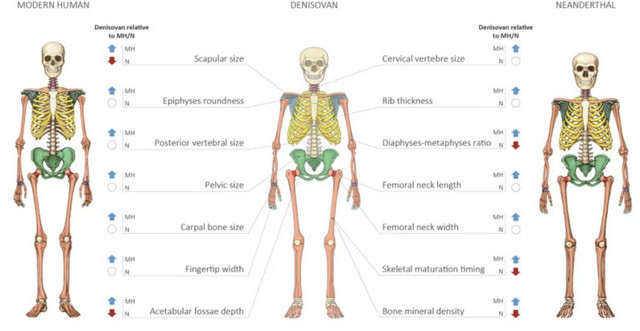
Kuvassa keskellä on tutkijoiden päätelmä denisovanihmisen luurangosta. Oikealla neandertalilainen, vasemmalla nykyihminen. Kuva: Maayan Harel / Hebrew University
Tutkimuksen tulokset on julkaistu Nature-sarjan Molecular Psychiatry -lehdessä. Samat tutkijat ovat kirjoittaneet useita artikkeleja ihmisen temperamentin, luonteenpiirteiden ja persoonallisuuden geneettisestä rakenteesta.
Koronavirus tuo samanlaisia ongelmia kuin esi-isillä
267 geenin evoluutiota vertaillaan tutkimuksessa simpanssien, nykyihmisen DNA:n ja Euraasiasta ja Euroopasta löydettyjen sukupuuttoon kuolleiden neandertalinihmisten muinais-DNA-löydösten välillä. Geenitutkimuksen kehitys avasi Lehtimäen mukaan ikkunan tutkijoille historiaan.
– Havaitsimme, että itsetuntemuksen säätelyverkossa simpanssit olivat alkeellisimpia, siinä välissä olivat neandertalinihmiset ja eniten näitä geenejä oli modernilla nykyihmisillä, Lehtimäki kertoo Ylelle.
Nykyihmiselle tehtiin tarkat psykologiset kokeet. Simpanssi on tutkimuksen mukaan 3-5-vuotiaan ihmisen tasolla. Neanderitalinihminen taas oli luovuudessa Lehtimäen mukaan simpanssin ja nykyihmisen välillä.
Sen jälkeen kun homo sapiensit muuttivat Afrikasta 65 000–55 000 vuotta sitten, he syrjäyttivät vähitellen neandertalinihmiset ja muut ihmislajit, jotka kaikki kuolivat sukupuuttoon noin 40 000 vuotta sitten.
– Tutkimusryhmä on kovasti innostunut, koska ajateltiin, että tämä tieto oli menetetty ikuisiksi ajoiksi historian hämärään. Se, että pystyimme löytämään nykyihmiselle nämä melkein tuhat geeniä, käänsi tilanteen.

Neandertalilaisten tekemä kädenjälki luolan seinässä Espanjassa. Kuva: H. Collado / Max-Planck- Gesellschaft
Luovuus ratkaisi selviytymisen
Nykyihmisille ominaisten geenien ansiosta voimme olla luovia taiteessa ja tieteessä, sosiaalisia ja elää pidempään olemalla aiempaa vastustuskyisempiä ikääntymistä, vammoja ja sairauksia vastaan.
– Tälläkin hetkellä kohtaamme koronapandemian vuoksi samanlaisia haasteita, joihin meidän on sopeuduttava luovasti, kuten esi-isämme oman aikansa haasteisiin, Lehtimäki sanoo.
Lehtimäen mukaan Homo sapiensin, neandertalinihmisten ja simpanssien proteiineja koodaavat geenit ovat lähes kaikki samanlaisia. Mutta eivät ihan samoja, ja se on nykyihmisen onni.
Geenit, jotka erottavat modernit ihmiset neandertalinihmisistä ja simpansseista, ovat tutkijoiden mukaan melkein kaikki RNA:sta valmistettuja säätelygeenejä, eivätkä DNA:sta valmistettuja proteiineja koodaavia geenejä.
– Proteiinia koodaavien geenien välissä on noin viisi kertaa enemmän pieniä säätelygeenejä, jotka eivät olekaan DNA:ta, vaan RNA:ta. Ne hienosäätävät kaikkia meidän oppimis- ja muistitapahtumiamme, aineenvaihduntaamme ja muuta. Se geenistön osa on ihmisellä erottava osa, Lehtimäki selittää.
Professori ei itsekään olisi uskonut, että asia voidaan selvittää. Samaa teknologiaa käyttäen on pystytty löytämään noin tuhat rasva-aineenvaihdunnan geeniä. Tätä tutkimusta ei Lehtimäen mukaan ole vielä julkaistu ja löydöksen merkityksen arviointi ihmisen evoluutioon on vielä kesken.
– Arvaus on, että se ei ole niin ratkaiseva kuin tämä löydös. Tämä jää pitkäksi aikaa henkilökohtaisen tutkimuksenurani huippulöydökseksi.
Lehtimäki muistuttaa, että geenit eivät määrää yksin käyttäytymistä, vaan voimme edelleen oppia ja sopeutua kokemuksista.
Lue lisää:
Tutkimus: Vakavia koronaoireita saavilla on ratkaisevassa kohdassa pätkä neandertalilaisten perimää
Ainutlaatuinen kuva sukualbumiin: Tältä näytti 70 000 vuotta sitten elänyt serkkumme
Geeneissämme piilottelee tuntemattomia esi-isiä – tutkijoiden mukaan niitä voi löytyä useita
Yhteistyö oli voimaa myös neandertalilaisille – ovela metsästäminen ei ole vain nykyihmisten taito
Totaalista hewonhuttua...

Liisa Keltikangas-Järvinen on hullumpi ja typerämpi puoskarihouru kuin olisin osannut ikinä kuvitellakaan...
Nykyihmisen ja neandertalilaisen sialiinimutaationjälkeiset ehdollistumiseen liittyvät aivokemikaalit poikkeavat yhden proteiinin TKTL1 osalta, joka poikkeaa YHDEN (1) emäsparin osalta näillä lajoilla. Tämä aiheuttaa sen, että nykyihmisellä on noin kaksinkertainen määrä mutta pienempiä aivosoluja kuin neandertalilaislla "serkuilla".
https://scitechdaily.com/key-differences-revealed-between-brains-of-modern-humans-and-neanderthals/
" Key Differences Revealed Between Brains of Modern Humans and Neanderthals
TOPICS: Brain Max Planck Institute Neanderthals Neuroscience Popular
By Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG) September 9, 2022

Scientists uncover a greater neuron production in the frontal lobe during brain development in modern humans than Neanderthals due to the change of a single amino acid in the protein TKTL1.
What makes modern humans unique? It is a question that has long been a driving force for researchers. Therefore, fascinating insights are revealed by comparisons with our closest relatives,the Neanderthals. The increase in brain size and increased neuron production during brain development are considered to be primary factors for the increased cognitive abilities that occurred during human evolution.
However, while both Neanderthals and modern humans develop brains of similar size, very little is known about whether modern human and Neanderthal brains may have differed in terms of their neuron production during development.
Scientists from the Max Planck Institute of Molecular Cell Biology and Genetics (MPI - CBG) in Dresden have discovered that the modern human variant of the protein TKTL1, which differs by only a single amino acid from the Neanderthal variant, in-creases a specific type of brain progenitor cells,called basal radial glia,in the modern human brain.Basal radial glial cells generate the majority of the neurons in the deve- loping neocortex, a part of the brain that is essential for many cognitive abilities.
Because TKTL1 activity is particularly high in the frontal lobe of the fetal human brain, the scientists conclude that this single human-specific amino acid substitution in TKTL1 underlies a greater neuron production in the developing frontal lobe of the neocortex in modern humans than in Neanderthals.
Just a small number of proteins have differences in the sequence of their amino acids – the building blocks of proteins – between modern humans and our extinct relatives, the Neanderthals and Denisovans. It is largely unknown what the biological significance of these differences is for the development of the modern human brain. In fact, modern humans and Neanderthals feature a brain, and notably a neocortex, of similar size, but whether this similar neocortex size implies a similar number of neurons remains unclear.

Microscopy picture of a dividing basal radial glial cell, a progenitor cell type that generates neurons during brain development. Modern human TKTL1, but not Neanderthal TKTL1, increa-ses basal radial glia and neuron abundance. Credit: Pinson et al., Science 2022 / MPI-CBG
The latest study of the research group of Wieland Huttner, one of the founding directors of the Max Planck Institute of Molecular Cell Biology and Genetics (MPI-CBG) in Dresden, addresses just this question. The research was carried out in collaboration with Svante Pääbo, director at the Max Planck Institute for Evolutionary Anthropology in Leipzig, and Pauline Wimberger of the University Hospital Dresden and their colleagues.
The scientists focus on one of these proteins that presents a single amino acid change in essentially all modern humans compared to Neanderthals, the protein transketolase-like 1 (TKTL1). Specifically, in modern humans, TKTL1 contains an arginine at the sequence position in question, whereas in Neanderthal TKTL1 it is the related amino acid lysine. In the fetal human neocortex, TKTL1 is found in neo-cortical progenitor cells, the cells from which all cortical neurons derive. Notably, the level of TKTL1 is highest in the progenitor cells of the frontal lobe.
New research reveals that a change in a single amino acid in the protein TKTL1 results in greater neuron production in the developing frontal lobe of the neocortex in modern humans compared with Neanderthals.
Anneline Pinson, the lead author of the study and researcher in Wieland Huttner’s group, set out to investigate the significance of this one amino acid change for neocortex development. Anneline and her colleagues introduced either the modern human or the Neanderthal variant of TKTL1 into the neocortex of mouse embryos.
They observed that basal radial glial cells,the type of neocortical progenitors thought to be the driving force for a bigger brain, increased with the modern human variant of TKTL1 but not with the Neanderthal variant. As a consequence, the brains of mouse embryos with the modern human TKTL1 contained more neurons.
More neurons in the frontal lobe of modern humans
After this, the investigators explored the relevance of these effects to human brain development. To this end, they replaced the arginine in modern human TKTL1 with the lysine characteristic of Neanderthal TKTL1, using human brain organoids. These are miniature organ-like structures that can be grown from human stem cells in cell culture dishes in the lab and mimic aspects of early human brain development.
“We found that with the Neanderthal-type of amino acid in TKTL1, fewer basal radial glial cells were produced than with the modern human-type and, as a consequence, also fewer neurons,” says Anneline Pinson. “This shows us that even though we do not know how many neurons the Neanderthal brain had,we can assume that modern humans have more neurons in the frontal lobe of the brain, where TKTL1 activity is highest, than Neanderthals.”
The researchers also discovered that modern human TKTL1 acts through changes in metabolism. Specifically, stimulation of the pentose phosphate pathway followed by increased fatty acid synthesis. In this way, modern human TKTL1 is thought to increase the synthesis of certain membrane lipids needed to generate the long process of basal radial glial cells that stimulates their proliferation and, therefore, increases neuron production.
“This study implies that the production of neurons in the neocortex during fetal deve-lopment is greater in modern humans than it was in Neanderthals, in particular in the frontal lobe,” summarizes Wieland Huttner, who supervised the study. “It is tempting to speculate that this promoted modern human cognitive abilities associated with the frontal lobe.”
Reference: “Human TKTL1 implies greater neurogenesis in frontal neocortex of modern humans than Neanderthals” by Anneline Pinson, Lei Xing, Takashi Namba, Nereo Kalebic, Jula Peters, Christina Eugster Oegema, Sofia Traikov, Katrin Reppe, Stephan Riesenberg, Tomislav Maricic, Razvan Derihaci, Pauline Wimberger, Svante Pääbo and Wieland B. Huttner, 9 Sepetember 2022, Science.
DOI: 10.1126/science.abl6422 "
TKTL1 and hominin cortical neurogenesis.

The single lysine-to-arginine substitution in modern human TKTL1 leads to greater bRG num-bers than in Neanderthals. These bRG in turn generate more neocortical neurons in modern hu-mans. Because TKTL1 expression in fetal human neocortex is particularly high in the developing frontal lobe, these findings imply that the frontal lobe of modern humans contains more neurons than that of Neanderthals.
" Neanderthal brain development
Neanderthal brains were similar in size to those of modern humans but differed in shape. What we cannot tell from fossils is how Neanderthal brains might have dif-fered in function or organization of brain layers such as the neocortex. Pinson et al. have now analyzed the effect of a single amino acid change in the transketolase-like 1 (TKTL1) protein on production of basal radial glia, the workhorses that generate much of the neocortex (see the Perspective by Malgrange and Nguyen). Modern humans differ from apes and Neanderthals by this single amino acid change. When placed in organoids or overexpressed in nonhuman brains, the human variant of TKTL1 drove more generation of neuroprogenitors than did the archaic variant. The authors suggest that the modern human has more neocortex to work with than the ancient Neanderthal did. — PJH
Structured Abstract
INTRODUCTION
The evolutionary expansion of the neocortex and the concomitant increase in neuron production are considered to be a basis for the increase in cognitive abilities that oc-curred during human evolution. Endocast analyses reveal that the endocranial vo-lume of modern humans and Neanderthals was similar, suggesting similar brain and neocortex size. But whether similar neocortex size implies similar neocortical neuron production remains unclear.
RATIONALE
Transketolase-like 1 (TKTL1) is a gene from the transketolase family that in fetal human neocortex is preferentially expressed in the two classes of neuroprogenitors, the apical progenitors in the ventricular zone and the basal progenitors in the sub-ventricular zone. The latter class of neuroprogenitors comprises two major types, the basal intermediate progenitors (bIPs) and the basal radial glia (bRG,also called outer radial glia). bRG exhibit cellular processes that promote their ability to self-amplify, and are the neuroprogenitor type considered to be a driver of the increase in cortical neuron production, which is a hallmark of the evolution of the human neocortex.
Reflecting their cell polarity, bRG undergo repeated asymmetric divisions that self-renew the bRG and generate one neuron each. Thereby, bRG generate more neurons over time than the other type of neuron-generating basal neuroprogenitors, the process-lacking bIPs whose neurogenic divisions are symmetric self-consuming.
TKTL1 is one of the few proteins with a single amino acid substitution found in essentially all present-day humans but absent from extinct archaic humans, the Neanderthals and Denisovans, and other primates. This human-specific amino acid substitution in TKTL1 is a lysine in apes and archaic humans but an arginine in mo- dern humans. We therefore investigated (i) whether TKTL1 has a role in neocortex development and affects neuroprogenitor numbers and (ii) whether both archaic TKTL1 (aTKTL1) and modern human TKTL1 (hTKTL1) exert similar effects on neuroprogenitors during neocortex development.
RESULTS
When expressed in mouse embryo neocortex, which lacks TKTL1 expression, hTKTL1 increased the abundance of bRG without affecting that of bIPs and that of apical progenitors. The effect of TKTL1 on bRG abundance was limited to hTKTL1; aTKTL1, which differs only by one amino acid, was unable to increase bRG abundance. The greater bRG abundance upon hTKTL1 expression resulted in an increase in cortical neuron production over time, specifically of the late-born upper-layer neurons rather than of the early-born deep-layer neurons. In the folded (gyren-cephalic) developing ferret neocortex, hTKTL1 expression increased not only bRG abundance but also the proportion of bRG with multiple processes, a hallmark of bRG that can self-amplify. As a consequence of this effect, gyrus size increased.
In fetal human neocortex, hTKTL1 was essential to maintain the full number of bRG, as CRISPR-Cas9–mediated hTKTL1 knockout in fetal human neocortical tissue re-duced this number. To further demonstrate the relevance of this effect, we converted hTKTL1 to the Neanderthal variant aTKTL1 in human embryonic stem cells and ge-nerated minibrain structures called cerebral organoids. The aTKTL1-expressing or-ganoids contained fewer bRG and neurons, hence the human-specific lysine-to-ar-ginine substitution in hTKTL1 is essential for maintaining the full number of bRG and neurons in this human brain model. In fetal human neocortex, hTKTL1 expression in neuroprogenitors increased during the course of neurogenesis and was particularly high in the developing frontal lobe as compared to the developing occipital lobe.
As to its mechanism of action, hTKTL1 increased bRG abundance via two metabolic pathways, the pentose phosphate pathway (PPP) followed by fatty acid synthesis. Inhibition of the PPP or of fatty acid synthesis, using a variety of specific inhibitors, completely suppressed the hTKTL1-induced increase in bRG abundance in embryo-nic mouse neocortex and reduced bRG numbers in fetal human neocortical tissue. This metabolic action of hTKTL1, but not aTKTL1, in bRG resulted in an increase in the concentration of acetyl–coenzyme A, the critical metabolite for fatty acid synthe-sis. Our data suggest that hTKTL1, but not aTKTL1, promotes the synthesis of mem-brane lipids containing a certain type of fatty acid that are required for the outgrowth of bRG processes and hence for the increase in bRG abundance.
CONCLUSION
In light of our finding that TKTL1 expression in fetal human neocortex is particularly high in the developing frontal lobe, our study implies that because of the single amino acid–based activity of hTKTL1, neocortical neurogenesis in modern humans was and is greater than it was in Neanderthals, in particular in the frontal lobe. "
https://www.mpi-cbg.de/research/researchgroups/currentgroups/wieland-huttner/research-focus
***
https://www.hs.fi/paakirjoitukset/art-2000007422953.html
https://psyk.fi/…/paivahoidosta-ja-opetuksesta-tunteita…/
Päivähoidosta ja opetuksesta tunteita herättävää tutkimusta
Kommentit