Ovatko "itämeresuomalaiset vallannet maansa kantapersermaaneilta"?
HAISTAPASKANTIETEILIJÖIDEN "RAISKAAJAKAANITEORIA" ON TÄYSIN UFOLOGINEN!
On huomattava, että tämä ei ole "perinteinen" hitleristis-hirohitoistinen pseudoteo-ria, sillä heidän tarkoituksensa EI ollut "vallata alempien rotujen naisia", vaan nimenomaan tappaa aivan erityisesti heidät pois "rotua pilaamasta"!
Se poikkeaa kuitenkin tuosta "toisesta" teoriasta VAIN "ARVOILTAAN", ei perusolet-tamuksiltaan "ihmisluonnosta" (jollaista ylihistoriallisena ei ole...). Se ei ole niiden vastainen, vaan se on NURINKÄÄNNETTY!
Se voidaan suit sait käden käänteessä kääntää taas alkuperäisin päin: "Kaanilaiset on tapetteva!"...
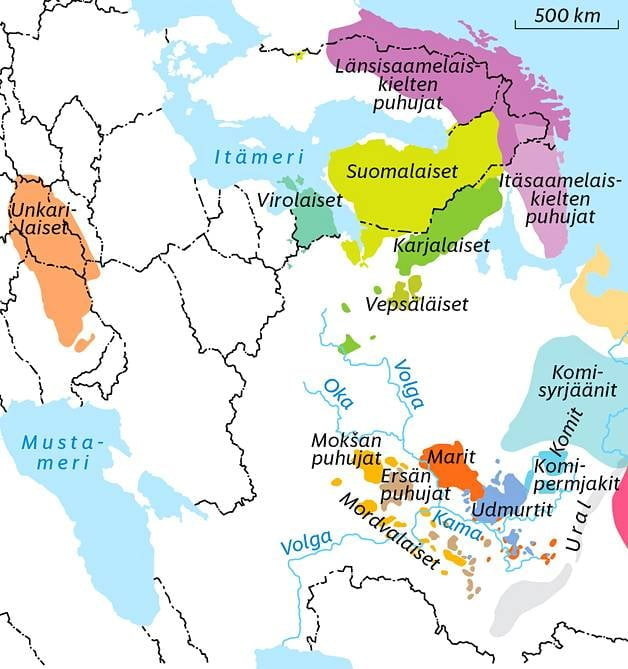
Genneettisltä isälinjoiltaan uralilaisempia kuin virolaiset (34%) latvialaisia (38%) ja liettualaisia (42%, suomalaiset 61%) näille tiedehuijareille ei ole lainkaan olemas-sa sen enempää kielellisesti kuin geneettisetikään; he ovat "sekundaryssiä" (mikä sikäläisten vaskistienkin kannattaisi pika pikaa noteerata...).
TÄSTÄ HAISTAPASKANTIETEEN HOURULALLATUKSESTA ON SAATANA TULTAVA LOPPU!!!

Itäinen perimä näkyy uralilaisia kieliä puhuvissa miehissä. (KUVA: Kimmo Taskinen / HS)
Kantagermaanit ovat todellinen muinainen kansa, joka on asunut luultavimmin keski-Euroopassa, ilmeisesti Tonavan alueella suurin piirtein meidän itäbalttilaisen (= voi-makkaasti SU-vaikutteisesti balttilaisen) vasarakirveskansamme aikalaisina. On luul-tavaa, että kanta- tai muutkaan germaanit eivät kuuluneet indoeurooppalaisten SO-TAVAUNUKANSOJEN kokoonpanoon, johon taas kuuluivat mm kelttiläis-romaaniset ja indo-iranilaiset heimot ja osa Volgan "vasarakirveskansasta" (mm. hevosjumalatar Ratainycia eli "Rattaiska" oli heidän jumaluutensa, ja IE-kielten kuuluisa sotavaunu-sanasto on ny-kyisissäkin balttikielissä VASARAKIRVESLAINOJA), ja niiden keltti-läis-romaniset ja slaavilaiset ja skandinaaviset germaanikielet sekä gootti vilisevät vasarakirveslainoja
Sen sijaan KANTAPERESERMAANIT on SEPITETTY "KANSA";joka olisi muinoin, ennen suomalaisugrilaisia "Kaanin-äpäröitä" muka asuttanut Viroa, Etelä-Suome ja Pohjois-Ruotsia.
https://hameemmias.vuodatus.net/lue/2014/09/yha-vaan-suomen-muinaiskielista
https://www.pirkanblogit.fi/2017/risto_koivula/suomessa-ei-ole-ollut-koskaan-kantapersermaaneja/
"Teorian" kehittäjä lienee tallinnalaisyntyinen Alfred Rosenberg, Saksan natsipuolu-een perustaja ja yksi ideologi, jolta Nürnbergissä putosi tuosta haistapaskanteorian tekeleestä aiheellisesti pää.
HM: Korjaus: Virolais-latvialainen "Affe" Rosenberg, jossa ei ollut pisaraakaan ker-maaniverta, ei keksinyt tuota ideaa omasta päästään, vaikka ns. maksoikin täyden hinnan. Toista tuomionsa perustetta, Saksan Kansallissosialistisen puolueen sijais-puheenjohtajan hommaa Hitlerin istuessa linnassa, hän oli hoitanut niin huonosti ja herättäen omien ja ulkomaisten aateveljien epäluottamusta mm. syntyperänsä takia (jonka mm. unkarilaiset tonkivat selville juurta jaksain), että siitä hyvästä hänelle olisi voitu myöntää vaikka rauhanpalkinto. Aatun palattua linnasta ruoriin ja julkaistua Mein Kampfin kannatus pomppasi prosentista-kahdesta 30:en tietämille. Hitler oli liittynyt Rosenbergin puolueeseen jolla oli jo silloin TUO ideologia, kuten myös armo- ton antisemitismi erästä toista kautta.Nämä eivät kuitenkaan vielä riitäneet tekemään fasisteista tappo-oppia jyrääviä natseja.Siihen "tarvittiin" myös erään Hans Bergerin "aivoteoriaa".
"Pohjolan kanta-arjalaisteorian (nimi tullee täällä saamen etelää tarkoittavasta sanasta oarje, joka on silloin tarkoittanut maanmuokkausta, -viljelyä, sitä harjoittavia naapuriheimoja, se on samaa balttilähtöä kuin iranin sana, syöttivät jo 1600-luvulla ruotsalaiset luterilaiset piispat saksalaiselle kuuluisalle fyysikolle ja matemaatikolle ja diplomaatille, fanaattiselle luterilaiselle monadologiselle (pitää muodollista logiikkaa "luonnon rakenneominaisuutena") filosofille Gottfried Wilhelm Leibnizille, joka julkaisi teoriakyhäelmän salanimellä Johannes Perignorus:
" Lukijan mielenkiintoa herättävät ensimmäisessä luvussa perusteelliset selvitykset germaanis-suomalaisten lainakosketusten varhaisimmasta tutkimuksesta. Saamme kokea tutkimuksen alkuvaiheet yhdessä Kustaa Aadolfin opettajan Johannes Bure-uksen, hampurilaisen lääkärin ja historioitsijan Martin Fogelin sekä turkulaisen laki-tieteen ja historian professorin Mikael Wexioniuksen kanssa. Tekijä oikaisee tässä Karstenin Georg Stiernhielmiä koskevan huomautuksen, jonka mukaan tämä olisi jo lausunut mielipiteensä lainasanojen suunnasta ja Skandinavian alkuperäisistä asukkaista.Todellisuudessa hän on esittänyt käsikirjoituksessaan Miscellania Etymologica »Finnonican» kohdalla vain joukon eri kielissä toisiaan vastaavia sanoja. Kiinnostus historiaa kohtaan on samoin seuraavien lainasanojen tutkijoiden johtolankana.
Myös tässä yhteydessä Kylstra voi täydentää aikaisempia tietoja.Erik Benzeliuksen - silloisen upsalalaisen kirjastonhoitajan - suhteet [Gottfried Wilhelm] Leibniziin olivat läheisemmät kuin Setälä, Wiklund ja Karsten luulivat.

Erik Benzelius nuorempi (1675–1743), Uppsalan arkkipiispa 1742-1943.

Gottfried Wilhelm Leibniz (1646 - 1716)
Voidaan todistaa, että Benzelius on käynyt Hannoverissa Leibnizia tapaamassa ja että Leibniz kirjoitti tämän ajatustenvaihdon pohjalta [salanimellä Johannes Dominicus Peregrinus] teoksen Dissertatio de origine Germanorum, jossa hän mainitsee Benzeliuksen kanssa Skandinavian alkuasukkaista käydyn keskustelun.

Tämä kysymys muodostui myöhemmän lainasanojen tutkimuksen ydinkohdaksi. "
Tuo kusetus on sivuluonne täällä Hesarin viikon valehistoriapierussa.
Mutta juuri se on kuitenkin yleisemmin se pääasia...
https://www.hs.fi/tiede/art-2000005912508.html
Suomalaisten juuret löytyivät Siperiasta – alkuperäisasukkaat toivat mukanaan kantauralin kielen
Yhteinen perimä kertoo, että esi-isämme toivat kielen tullessaan ja iskostivat sen uu-sille alueille. Yhteinen perimäaines puhuu sen puolesta, että alkuperäinen kantaura-lin kieli levisi ja haarautui ihmisten mukana. Samaa todistaa se tutkijoiden huomio, jonka mukaan uralilaiskansojen geneettiset erot ovat sitä suurempia, mitä suurempia ovat kielelliset erot.
... Kieli voi levitä myös lainana niin, että ihmisyhteisöt omaksuvat jonkin toisen yhteisön kielen ja luopuvat entisestä. Tästä ei voi olla kyse uralilaisten kielten leviämisessä. Niiden puhujat todella veivät kielensä uusille asuinsijoilleen.
Poikkeuksen tutkimuksessa muodostavat nykyiset unkarilaiset, kaikista suurin urali-laisten kielten puhujien ryhmä. Uralilaisia kieliä puhuvia ihmisiä on 20–25 miljoonaa, joista Unkarissa asuu tätä nykyä kymmenisen miljoonaa.
”Nykyiset unkarilaiset muistuttavat geneettisesti eniten lähinaapureitaan Itä-Euroo-passa eivätkä jaa paljoakaan geenejä muiden uralilaisten kansojen kanssa. Heiltä puuttuvat siperialaiset ainekset,jotka muilla uralilaisilla kansoilla on”, kertoo tutkimus-ta johtanut Kristiina Tambets Tarton yliopiston perimäntutkimuksen instituutista.
Kiinnostavaa on,että vaikka unkarilaisilla ei juuri ole yhteisiä geneettisiä erityispiirtei-tä kieliperheen muiden jäsenten kanssa, he puhuvat yhä uralilaista kieltä. Uralilaiset saivat siis istutettua kielensä hallitsevaan asemaan uusilla asuinalueillaan itäisessä Keski-Euroopassa, vaikka heidän geneettinen jälkensä lopulta katosi.
Kyse voi olla siitä, että muuttajat edustivat tavalla tai toisella hallitsevaa kulttuuria. "
K: Unkarilaiset tulivat nykyisille alueilleen hunnien mukana tai perässä 300-luvulla j.a.a. He tulivat idempää kuin ensimmäinen eurooppalainen kansa, jonka kimppuun he iskivät, eli Donin alueella vaikuttaneet iranilaiskieliset alaanit. Alaaneista länteen olivat germaaniset, vahvasti vasarakirves- ja sen mukana myös jossakin määrin SU-vaikutteiset (itä)gootit.
TÄTÄ SAMAA AJETAAN NYT KUIN KÄÄRMETTÄ PIIPPUUN MYÖS MUKA "ITÄ-MERENSUOMALSITEN JA SAAMELAISTEN TAUSTANA" - ei ehkä juuri hunneja, mutta joitakuita muita...
HS: ”Muuttajaryhmä ei ollut välttämättä kovin suuri, mutta heillä oli arvovaltaa ja heidän kieltään alettiin puhua seuraavissa sukupolvissa”, Tambets selittää.
K. : Unkarilaset tulivat idästä - mutta he olivat menneet sinne "lännestä", Volgan alueelta, mahdollisesti vasarakirveskansan ja skyyttien "tieltä" - jotkut muut kuten Suomen vasarakirveskansa olivat silloin lähteneet länteen. On siis syytä olettaa, että unkarilaiset eivät olleet silloinkaan mitään "umpi-itäisiä" - koska sitä eivät olleet itse hunnitkaan, vaikka he olivat tuleet Kiinasta.
HS: Tutkimuksen tekoon osallistunut Outi Vesakoski Turun yliopiston biologiaa ja kielihistoriaa yhdistävästä hankkeesta sanoo, että Unkarin uralilaiset ovat ilmeisesti olleet ratsastajakansaa.
”Lainasanojen perusteella he ovat vaellelleet edestakaisin nykyisen uralilaisalueen eteläpuolella. Heidän siirtymisensä Keski-Eurooppaan on hyvin dokumentoitu. Siellä he onnistuivat tekemään niin suuren vaikutuksen, että heidän kielensä jäi sinne pysyvästi”, Vesakoski sanoo.
Vaikka nykyunkarilaisten perimästä ei löytynyt itäisiä aineksia, kuva voi muuttua, jos pystytään saamaan dna:ta muinaistutkijoiden kaivauksilta. Kaiken järjen mukaan Unkarin alueen muinais-dna:ssa voisi näkyä geneettisiä yhteyksiä kielisukulaisiin.

HS: " Pohjoisempana Itämeren rannikolla uralilaiset saivat niin ikään kielensä iskos-tettua uusille alueille. Täällä suomen, viron, vepsän ja karjalan kieli ovat melko uusia tulokkaita.
”Nykykäsityksen mukaan itämerensuomalaiskielten esimuodot tulivat hieman ennen ajanlaskun alkua Itämeren rannoille. Ihmisiä täällä on asunut tuhansia vuosia kauemmin,mutta emme tiedä,mitä kieltä he puhuivat”,Tambets kertoo. "
K.: HEWOWWITTUA!
Suomen kampakeraaminen kansa on 7000 vuotta sitten tuonut tänne SU-kielen, joka on ollut nykykielistä lähempänä unkaria kuin suomea.
He ovat tuoneet myös tattarinviljelyksen - 1000 vuotta ennen kuin trattbägarit - kantagrmaanien kaverit sittemmin - vehnänviljelyksen Ruotsiin!
Juuri he toivat N1c-haplon, kampakeramiikan, ja maalasivat Astuvansalmen kalliomaalaukset:
http://www.helsinki.fi/hum/ajankohtaista/2013/01/0128b.htm
" Maanviljely levisi Suomeen Itä-Aasiasta jo 7000 vuotta sitten
Siitepölytutkimusta ja arkeologiaa yhdistävä tutkimushanke "Kouvolan seutu muinai-suudessa, korpea vai kaskenkaatajia" on selvittänyt vuosina 2010-12 nykyisen Kou-volan pohjoisosan esihistoriaa ja maankäyttöä jääkaudelta 1200-luvulle jKr. Hank-keen tuloksena on syntymässä uudenlainen näkemys suomalaisen maatalouden alkutaipaleesta.
Tutkimusalue sijaitsee Kouvolan pohjoisosassa,Jaalan Huhdasjärvellä.Alueen maa-talousasutuksen on aiemmin oletettu syntyneen vasta historiallisella ajalla, viimeis-tään 1500-luvulla. Arkeologiset löydöt kuitenkin osoittavat, että alueella on elänyt yhteisö,joka on ylläpitänyt kalmistoa 600-luvulta 1100-luvulle jKr.Tutkimuksen ensim-mäisenä tavoitteena oli selvittää, harjoittiko yhteisö maa- vai erätaloutta. Toisena kysymyksenä olivat maanviljelyn alkuvaiheet - onko Huhdasjärveltä löydettävissä yhtä vanhoja maanviljelyn tuloksia kuin läheltä, Repoveden kansallispuiston alueelta muutama vuosi sitten löydettiin.
Saadut tutkimustulokset muuttavat käsitystä maanviljelyn alkuvaiheista niin kivikau-den kuin rautakauden osalta. Varhaisin metsänraivausvaihe ajoittuu kivikaudelle 5300 - 4000 eKr. Ensimmäisenä viljelykasvina oli tattari 5300 eKr., ja noin 4200 eKr. ilmestyvät ensimmäiset merkit ohran viljelystä. Nämä viljelyn merkit ajoittuvat kivikaudelle, samaan aikaan saviastioiden valmistustaidon omaksumisen kanssa (varhaiskampakeramiikka). "
HS: " Se tiedetään, että nykyisen Suomen alueelle uralilaiset kielet olivat jo aiemmin tulleet saamelaiskielinä, mutta Suomessa oli väestöä jo ennen saamelaiskieltenkin puhujia. "
Tuosta ei ole mitää näyttöä!
Saamelaikielet eivät ole olennaisesti vanhempia kuin itämerensuomalaisetkaan!
Molemmilla on sama alkuperä kampakeraamisesta ja vasarakirveskielstä, mutta ilmeisimmin eri paikoissa!

Haistapaskantiedesyöttiläät pällistelee...
WITTUUN SAATANAN RIAHU "HYVÄN SÄÄN AIKANA"!!!!
" HS: " Itämerensuomen kantamuodon tuojat saapuivat Volgan alueelta ilmeisesti reittiä, joka seuraili Daugava-jokea nykyisessä Latviassa. Näin todistelee Tarton yliopiston arkeologian professori Valter Lang, joka on julkaissut tänä vuonna kirjan Läänemeresoome tulemised eli Itämerensuomen saapuminen.
Helsingin yliopiston itämerensuomalaisten kielten professori Riho Grünthal kuvaa kirjaa arviossaan Elo-lehden arviossaan käänteentekeväksi.
Grünthalin mukaan Langin tulkinta Suomen kielihistoriasta on uusi ja vallankumouksellinen. "
”Kyse ei ollut germaanisia kieliä puhuvan väestön kulkeutumisesta kantasuomen alueella, vaan päinvastaisesta prosessista!”
K.: "... Eli että "itämerensuomalaiset valtasivet Hitlerin (olemattomien) kanta-arjalaisten alueita" ... pitäisiköhän vielä "PALAUTTAAKIN"...?
Haistapaskantieteilijät ovat valtaamassa myös Tartton yliopistoa... Riehu järjestää...
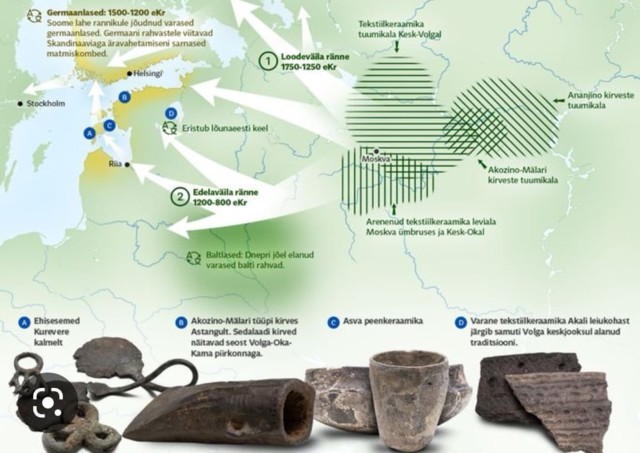
HS: " Suomeen suomen kielen kantamuoto on Langin mukaan saapunut Virosta roo-malaisella rautakaudella. Se ulottuu vuodesta 500 ennen ajanlaskun alkua vuoteen 400 jälkeen ajanlaskun alun. "
K.: Hewowwittua!!!
Itämerensuomalainen kielimuoto ON SYNTYNYT TÄÄLLÄ - balttilaisen vasara- kirveskielen vaikuttaessa (suomalais)ugrilaiseen kampakeraamiseen kieleen.
Kantapersermaaneita ei näkynyt eikä kuulunut PIERUNHAJUAKAAN mailla eikä halmeilla!!!
HS " Kantasuomen puhujat levittäytyivät Suomenlahden ylitse Varsinais-Suomen alueelle.
Virossa ja Etelä-Suomessa on saatettu puhua monia eri kieliä ennen uralilaisten tuloa, muun muassa esigermaania ja nyt jo hävinneitä muinaiskieliä. Hallitsevaksi kieleksi muodostui lopulta kuitenkin idästä tulleiden siirtolaisten puheenparsi. "
K.: TUOLLAISISTA EI OLE ENSIMMÄISTÄKÄÄN NÄYTTÖÄ - JOTEN SELLAISA EI OPIDÄ OLETTAAKAAN!!!!
HS: " Langin ajatusta kantasuomen saapumisesta tukee isälinjassa periytyvä Y-kro-mosomin haploryhmä, joka näyttää kulkeutuneen idästä länteen samaa reittiä kuin kielikin. "
HEWOWWITTUA SE SIIHEN AIKAAN VASTA OLE KULKEUTUNUT!!!!
Kampakeraaminen kulttuuri on iät ja ajat tiedetty N-haplo-valtaiseksi!!!
HS: " Volgalta Keski-Venäjältä on pitkä matka Suomenlahdelle. Uralilaisten kielten puhujien asuinalueet muodostavat karttakuvassa Itämerelle päin lähes yhtenäisen alueen. Ketjun katkaisee kuitenkin suuri valkoinen alue Karjalan itäpuolella ja Moskovan pohjoispuolella.
Valkoisella alueella ihmiset puhuvat venäjää, mutta he ovat silti uralilaiskielten puhujien sukulaisia, vahvistaa Tambetsin ja kumppanien tutkimus.
Pohjois-Venäjältä saaduista geeninäytteistä löytyi samaa siperialaista ainesta, joka yhdistää suomen sukukielten puhujia.
”He todella ovat itämerensuomalaisten kielten puhujien sukulaisia, koska he geneettiseltä profiililtaan muistuttavat näitä”, Vesakoski selittää.
Kielihistoriasta on arveltu, että siellä on tapahtunut kielenvaihto venäjään väestöpohjan pysyessä entisellään.
Muun muassa sikäläiset paikannimet viittaavat siihen, että siellä puhuttiin alle tuhat vuotta sitten länsiuralilaisia kieliä. "
K.: Ihan vapaasti...

Valter lang, uusi alfredrosenbergilainen hölynpölypuoskaritieteilijä.
HS: " Uralilaisessa perimässä näkyy myös sukupuoliero. Itäinen yhteys on paljon vahvempi miehissä kuin naisissa.
Ural-vuorten länsipuolella asuvilla uralilaisten kielten puhujilla on Länsi-Siperiaan viittaava Y-kromosomilinja, joka löytyy puolelta suomalaisista ja saamelaisista miehistä sekä 40 prosentilta virolaisista miehistä.
”Tätä ei löydy äitilinjoista. Ne ovat samanlaisia kuin muillakin eurooppalaisilla”, Tambets kertoo.
Geenit näyttäisivät antavan kuvan, että uralilaiset kielet toivat idästä miehet, jotka sitten ottivat vaimoikseen paikallisia naisia. "
K.: Näin tapahtui varmaan silloin 7000 vuotta sitten...
Tosin SU-kansojen naapurit eivät varmaan ole kovin palhjon silloisten alkuperäisten naisten heimojen jälkeläisiä...
Naiset ovat vaan vaihtuneet enemmän kuin miehet.
Myös latvialasissa ja liettualaisissa näkyy eritääin vahva uralilainen vaikustu nimenomaan miehissä.
HS: ”Ei voi sanoa, että vain miehiä tuli, mutta he jättivät selvimmät merkit itsestään”, Tambets sanoo.
Yhden Y-kromosomin hallitseva asema perimäaineistossa näyttää kertovan myös kieltä levittäneiden yhteisöjen sosiaalisesta rakenteesta.
”Tämä kromosomin hallinta jatkui useamman kuin yhden sukupolven ajan. Se osoit-taa, että sen aikaisessa yhteiskunnassa oli sosiaalisia tekijöitä, joiden ansiosta kro-mosomia kantavat miehet onnistuivat lisääntymään paremmin kuin muut”, Tambets kertoo.
Sosiaalinen rakenne saattoi esimerkiksi olla sellainen,että jonkin tietyn suvun miehet olivat johtavassa roolissa ja saivat siksi eniten jälkeläisiä. Tämä on vain arvailua, sillä geeneistä sitä ei voi pitävästi päätellä.
Myös Lang hyväksyy ajatuksen, että uralilaisia kieliä länteen levittäneet yhteisöt olivat miesvaltaisia. Se ei vielä kerro miesten välisistä hierarkioista.
Suomen kielen esimuotojen saapumisen ajallisista vaiheista geenien ja kielen haarautumista käsittelevä tutkimus ei kerro mitään.
Ennestään tiedetään kielitieteellisen päättelyn perusteella,että kantauralin ensimmäi- nen jakautuminen suomalais-ugrilaisten ja samojedikielten välillä tapahtui 6000 – 4000 vuotta sitten. "
K.: Se tapahtui jo 7000 vuotta sitten, kun jotkut lähtivät Suomeen!

Ensimmäiset kampakerramiset esi-isämme ovat saattaneet olla vaikkapa tämän näköisiä.
(Korjaus: Näillä on yleisin intiaanihaploryhmä Q. Heidän kielensäkin on sukua intiaa- nien Na Dene -kielikunnalle.Erään neuvostoliittolaisen lähteen mukaan vaan he ovat "antropologisesti identtisiä" naapureidensa hantien kanssa. Tämä oli vanhentunutta tietoa. Nämä ovat tulleet myöhemmin alueelle etelästä. HM)
HS: Arvio itämerensuomalaisten kielten saapumisesta tänne pari tuhatta vuotta sitten perustuu arkeologisiin ja kielitieteellisiin todisteisiin.
Uusi Genome Biology -lehdessä julkaistu tutkimus yhdisti geeniaineistot uralilaisten kielten sanastotietokantaan, jonka ovat koonneet ja julkaisseet Turun ja Tampereen yliopiston tutkijat. Tietokannassa on annettu numeerinen arvo perussanojen välisille etäisyyksille eri uralilaiskielten välillä.
”Perussanat ovat niitä, jotka esiintyvät kulttuurista riippumatta. Jokaisessa kielessä on sanat kuu, vesi, ruoka,äiti ja maha. Lumi ja korppikotka taas eivät ole perussa-nastoa, koska niiden tarkoittamia asioita ei ole kaikkialla”, Vesakoski selittää.
Tietokannassa samaa alkuperää olevat sanat kuten suomen ja viron sanat kuu saa-vat arvoksi ykkösen, kun taas eri lähteistä juontavat äiti ja ema antavat arvon nolla.
”Aineistosta saa tehtyä puumallin kielten erkaantumisesta. Puumallista saadaan mitattua eri kielten etäisyydet toisistaan: kuinka erilainen suomi on virosta, pohjois-saamesta tai komisyrjäänistä”, Vesakoski kertoo.
Tutkijat osoittivat, että geneettisiin etäisyyksiin perustuva uralilaisten kansojen sukupuu korreloi selvästi kielellisen sukupuun kanssa.
Tambets muistuttaa, että vaikka uralilaisten kielisukulaisuus on selvä, yhteinen perimäosuus on kuitenkin pieni.
”Ihmiset ovat kaikkein lähintä sukua maantieteellisille naapureilleen. Sen näkee jo ulkonäöstä. Siperiassa asuvat uralilaisten kielten puhujat muistuttavat muita Siperian asukkaita ja Euroopassa asuvat muita eurooppalaisia”, Tambets sanoo.
Pieni osa perimää kertoo kuitenkin kiehtovaa tarinaa kielten puhujien yhteisistä juurista ja historiasta. "
Haploryhmä N1 on ARKTINEN, ei välttämättä sen "itäaasialaisempi" kuin muutkaan europidit
Jos se sitten kuitenkin ON itäaasialsien , niin "so what"?
Se on ollut vallitseva haplorymä pohjoisessa Euraasiaas 7000 vuotta.

https://www.eupedia.com/europe/Haplogroup_N1c_Y-DNA.shtml
Haplogroup N1c is found chiefly in north-eastern Europe, particularly in Finland (61%), Lapland (53%), Estonia (34%), Latvia (38%), Lithuania (42%) and northern Russia (30%), and to a lower extent also in central Russia (15%), Belarus (10%), eastern Ukraine (9%), Sweden (7%), Poland (4%) and Turkey (4%). N1c is also pro-minent among the Uralic speaking ethnicities of the Volga-Ural region, including the Udmurts (67%), Komi (51%), Mari (50%) and Mordvins (20%), but also among their Turkic neighbours like the Chuvashs (28%),Volga Tatars (21%) and Bashkirs (17%), as well as the Nogais (9%) of southern Russia.
N1c represents the western extent of haplogroup N, which is found all over the Far East (China, Korea, Japan), Mongolia and Siberia, especially among Uralic speakers of northern Siberia. Haplogroup N1 reaches a maximum frequency of approximately 95% in the Nenets (40% N1c and 57% N1b) and Nganassans (all N1b), two Uralic tribes of central-northern Siberia, and 90% among the Yakuts (all N1c), a Turkic people who live mainly in the Sakha (Yakutia) Republic in central-eastern Siberia.
Distribution of haplogroup N1c in Europe

Phylogeny of N1c
If you are new to genetic genealogy, please check our Introduction to phylogenetics to understand how to read a phylogenetic tree.
Note that N1c was known as N3 and N1c1 as N3a in the official phylogeny prior to 2008.
N1c (L729)
N1c1 (M46/Page70/Tat)N1c1a (M178): found in Siberia (Khakass-Daurs) N1c1a1 (L708): found in Siberia (Anayins)N1c1a1a (P298): found in Siberia (Yakuts) N1c1a1a1 (L392, L1026): Finno-Ugric branch; found throughout north-east Europe N1c1a1a1a (CTS2929/VL29): Baltic-Finnic branchN1c1a1a1a1 (L550): West Finnic branch; found around the Baltic Sea and in places settled by the Vikings N1c1a1a1a1a (L1025)
N1c1a1a1a1a1 (M2783): found especially in Balto-Slavic countries, with a peak in Lithuania and Latvia
N1c1a1a1a1a2 (Y4706): found mostly in Finland and Scandinavia
N1c1a1a1a2 (CTS9976): East Finnic branch; found among the Chudes (Karelia, Estonia)
N1c1a1a1a2a (L1022)
N1c1a1a1a2a1 (Z1936): Finno-Permic branch; found in the Volga-Ural region and among the Karelians and SavoniansN1c1a1a1a2a1a (Z1925): found in Finland, Lapland, Scandinavia, the Volga-Ural and the AltaiN1c1a1a1a2a1a1 (Z1933)
N1c1a1a1a2a1a1a (Z1927): found among the Karelians
N1c1a1a1a2a1a1b (CTS8565): found among the Savonians
N1c1a1a2b (L1034): Ugric branch; found in and around Hungary and in Central Asia (Kazakhstan)
N1c2 (L666): found in Russia
N1c2a (M128)
N1c2b (P43): found in the Volga-Ural region
N1c2b1 (P63)
N1c2b2 (L665)
Totuuteen pyrkivienkään eurotieteilijöiden kaaliin ei mahdu, että väestö tai kultturi voisi koskaan levitä pohjoisesta etelään
Origins & History
Haplogroup N is a descendant of East Asian macro-haplogroup NO. It is believed to have originated in Indochina or southern China approximately 15,000 to 20,000 years ago.
Haplogroup N1* and N1c were both found at high frequency (26 out of 70 samples, or 37%) in Neolithic and Bronze Age remains (4500-700 BCE) from the West Liao River valley in Northeast China (Manchuria) by Yinqiu Cui et al. (2013). Among the Neolithic samples, haplogroup N1 made up two thirds of the samples from the Hong-shan culture (4700-2900 BCE) and all the samples from the Xiaoheyan culture (3000 - 2200 BCE), hinting that N1 people played a major role in the diffusion of the Neo-lithic lifestyle around Northeast China,and probably also to Mongolia and Siberia.
Ye Zhang et al. 2016 found 100% of Y-DNA N out of 17 samples from the Xueshan culture (Jiangjialiang site) dating from 3600–2900 BCE, and among those 41% belonged to N1c1-Tat. It is therefore extremely likely that the N1c1 subclade found in Europe today has its roots in the Chinese Neolithic. It would have progressively spread across Siberia until north-eastern Europe, possibly reaching the Volga-Ural region around 5500 to 4500 BCE with the Kama culture (5300-3300 BCE), and the eastern Baltic with the Comb Ceramic culture (4200-2000 BCE), the presumed ancestral culture of Proto-Finnic and pre-Baltic people. "
HM: Ei ole selvää, että N1c-haplo olisi Kiinasta: se on aivan hyvin voinut myös mennä pohjoisilta alueilta sinne eikä päinvastoin!
Uusin tutkimus antaa tähän yllärin:
KIINALAISET OVAT ITSEKIN ARKTISTA ALKUPERÄÄ!
https://www.tiede.fi/comment/2437085#comment-2437085
" Muinaiset beringiläiset olivat "kiinalaisia" - tai sitten kiinalaiset ovat muinaisia beringiläisiä!
https://www.nytimes.com/2018/01/03/science/native-americans-beringia-siberia.html?smid=fb-share
In the Bones of a Buried Child, Signs of a Massive Human Migration to the Americas
... " Beringia refers to Alaska and the eastern tip of Siberia, and to the land bridge that joined them during the last ice age. Appearing and disappearing over the eons, it has long been suspected as the route that humans took from Asia to the Western Hemisphere.
There has been little archaeological evidence,however,perhaps because early coas- tal settlements were submerged by rising seas. Thanks to her unique position in the Native American family tree, Xach’itee’aanenh T’eede Gaay has given scientists a clear idea how this enormous step in human history may have happened.
Her ancestors - and those of all Native Americans - started out in Asia and share a distant ancestry with Chinese people. In the new study, the scientists estimate those two lineages split about 36,000 years ago.
The population that would give rise to Native Americans originated somewhere in northeast Siberia, Dr.Willerslev believes.Archaeological evidence suggests they may have hunted for woolly rhino and other big game that ranged over the grasslands."
" There is little evidence of agriculture or domesticated animals in Siberia during the Neolithic,but pottery was widely used. In that regard it was the opposite develop-ment from the Near East,which first developed agriculture then only pottery from cir-ca 5500 BCE, perhaps through contact with East Asians via Siberia or Central Asia.
Mazurkevich et al. 2014 confirmed the presence of N1c in the Comb Ceramic cul-ture with a sample from the Late Neolithic site of Serteya II in the Smolensk region of Russia, near the Belarussian border, which dates from the middle of 3rd millenium BCE.
https://www.academia.edu/18216897/The_first_results_of_genetic_typing_of_local_population_and_ancient_human_bones_in_upper_dvina_region" Published in Archaeology of lake settlements IV-II mill. BC: chronology of cultures, environment and paleoclimatic rhythms. Saint-Petersburg, 2014. Mazurkevich A., Polkovnikova M., Dolbunova E. ed. P. 287 -294. 290
THE FIRST RESULTS OF GENETIC TYPING OF LOCAL POPULATION AND ANCIENT HUMAN BONESIN UPPER DVINA REGION Chekunova E.M. 1, Yartseva N.V. 1, Chekunov M.K. 2, Mazurkevich A.N.



The merger of the two groups, Indo-European R1a and Proto-Uralic N1c1, gave rise to the hybrid Kiukainen culture (2300-1500 BCE).
Modern Baltic people have a roughly equal proportion of haplogroup N1c1 and R1a, resulting from this merger of Proto-Uralic and Northeast Indo-European populations.
Tässä nähdään tilanne, kun kampakeraaminen Kaman kulttuuri, N1c, saapui mm. Astuvansalmelle 7000 vuotta sitten.
Yksinkertaisin selitys on paras?
Vaikka lähes kaikki tuunetut naisvaltaiset kulttuurit (kuten jukagirit) kuuluvat arktisiin kansoihin,niin yksikertaisin selitys paleoliittiselta kaudelta periytyvälle vallitsevalle isälinjalle on asutushistorialtaan jatkuva, "miesvaltainen" (patrilokaalinen) yhtei-sö, jossa naiset vaihtavat ryhmää ja asuinseutua paljon enemmän kuin miehet.
Tässä ei ole mitään "miessovinistista" eikä "patriarkalistista",sillä samaan aikaan eri- näiset merkit saatavt viitata siihen, että Suomen itäbalttilainen vasarakirveskulttuuri, toisin kuin sotavaunukansan osana ollut vasarakirveskulttuuri, olisi ollut naisvaltaista!
Se oli monissa suhteissa korkeatasoisempaa kuin kampakeraaminen kulttuuri, erityisesti maanviljelyksessä, mutta ei välttämättä kaikissa suhteissa.
Tällaisia seikkoja ovat:
- vasarakiveskansa ei näytä heimona olleen missään poliittisessa vallassa muiden yli, ei "herrakansaa" vaan raivaajakansaa, vaikka se on tullut myöhemmin ja kulttuu-risesti kehittyneemmältä alueelta.Sen sotiminen ja tehokas aseistus oli puolustuksel- lista (ennen kuin keksittiin sotavaunut, mihn Suomen vasarakirveskansa ei osallistu-nut, toisin kuin Volgan; vasarakirveskansa piti yli kaiken kiinni muokkaamistaan maa- ja vesialueista.
- vasarakirveskansan keramiikka on keittiötyövälinepyyppistä, kun taas kampake-ramiikka on varastointi- ja kuljetustyyppistä: mm. suurikoisempaa. Ero pikemminkin sen kun kasvoi vaan, ja molemmat ehkä käyttivät molempien keramiikkaa.
-vasarakirveskansa näyttäisi menneen Suomessa erityksissä "omista" jossakin mää- rin "takapakkia" joissakin miehisiksi mielletyissä suhteissa: esimerkiksi komeimmat rakennukset ovat vanhimmalta ajalta ja silloisen kultturipiirin itäisimmiltä alueilta.
-patrilokaalinen miesvaltainen naapurusto,ja liikkuva väestö (saamelaiset?) vain vah- vistaa tätä mahdollisuutta, sillä naapuriheimot tekivät heimoyhteiskunnassa useinkin tasan päin vastoin erilaissa perusratkaisuissa,kuten perhesysteemissä.Heimot myös jakautuivat usein jonkin peruserimielisyyden takia, joka sitten jatkui ja jatkui heimo-erona, ja saatettiin tehdä yhteistyössä molempia ratkaisuja, jos uudestaan taas ystävystyttin.
https://www.sciencedirect.com/science/article/abs/pii/S2352409X21004314?via%3Dihub
The palaeoenvironment and settlement history of a lakeshore setting: An interdisciplinary study from the multi-layered archaeological site of Serteya II, Western Russia
The Bronze Age Indo-European Fatyanovo-Balanovo culture (3200-2300 BCE) progressively took over the Baltic region and southern Finland from 2500 BCE (see History of haplogroup R1a).
... "
https://yle.fi/uutiset/3-11546935
Kalliomaalauksista saatiin uutta tietoa Nasan käyttämällä tekniikalla – voiko kallioon kuvattu hahmo olla saamelaisjumalattaren esiäiti?
Alempana jutussa olevista kuvista pääset katsomaan, miten selvästi tuhansia vuosia vanhat hahmot paljastuvat kalliosta.
17.9.2020 klo 18.05

Mikkelin Astuvansalmesta löytyi tämä hahmo, joka vasemmalla näkyy himmeällä alkuperäisessä värissään. Pekka Honkakoski on tulkinnut hahmon muistuttavan saamelaista jousikätistä jumalatarta, Juksáhkkáá.
Kun iisalmelainen lääkärinä työskentelevä Pekka Honkakoski sai muutama vuosi sitten käyttöönsä Nasan käyttämää tekniikkaa, alkoivat kalliomaalausten tarinat aueta aivan uudella tavalla.
Valokuvausta harrastava Honkakoski käytti kalliomaalausten tutkimiseen paitsi tätä arkeologeillekin tuttua, satelliittikuvauksissa käytettyä menetelmää, mutta myös eri-tyistä valotustekniikkaa. Näin kallioista alkoi paljastua uusia tarinoita paljon aiempaa enemmän.
Kun kalliomaalauksia löydettiin näin runsaasti, antoi se sijaa aivan uudenlaisille tulkinnoille.

https://images.cdn.yle.fi/image/upload//w_1198,h_675,f_auto,fl_lossy,q_auto:eco/13-3-11549686.jpg
{kind=link}
Liu'uttamalla pystyt havaitsemaan, kuinka värivarianssimuutoksella kalliomaalaukset tulevat vahvemmin esiin, jolloin niiden tulkitseminen helpottuu. Toinen kuvista on alkuperäisin värein, toisessa värivarianssimuutos. Kuvan hahmon Honkakoski on tulkinnut muistuttavan Juksáhkká-jumalatarta. Kuvat: Pekka Honkakoski
Pohjoismaiden laajimmat ja vanhimmat ihmisen tekemät kalliomaalaukset ovat 3500 – 7000 vuotta vanhoja. Niitä voi löytää esimerkiksi Suomussalmelta, Hossan Värikal-liolta sekä Mikkelin Astuvansalmelta. Näiden seutujen kalliomaalauksista on paljas-tunut teoksia, joita uudenlaisen tekniikan avulla on päästy tulkitsemaan paremmin.
Maalausten hahmoissa ja kuvastoissa voi nähdä samankaltaisuuksia esimerkiksi saamelaisten rumpujen kuviin.
Lue myös: Tutkimusmatkailija löysi Hossasta lisää kalliomaalauksia – hyödynsi NASAn menetelmiä
Voisiko kuvassa olla jumalatar Juksáhkkán esivanhempi?
Kalliomaalauksia tulkitessa kannattaa pitää mielessä,että kyseessä on tuhansia vuo- sia vanhoja teoksia aivan erilaisen kulttuurin aikakaudelta. Tulkinnoissa on Honka-kosken mukaan pyöräytettävä omia ajatuksiaan lapsen ajattelutapaan, etteivät nykyiset tiedot ja ajatukset liikaa vääristäisi tulkintoja.
Honkakoski on pitkän linjan valokuvausharrastaja, joka on erityisen kiinnostunut harvinaisemmista kuvauskohteista, niin lumihiutaileista kuin juuri kalliomaalauksista. Honkakoski on omien sanojensa mukaan tutkimusmatkailija ja häntä kiehtovat usein sellaiset paikat, joihin ihan jokainen valokuvaaja ei välttämättä päädy.
Honkakoski havaitsi löytämissään maalauksissa hahmon,joka muistuttaa saamelais- rummuissa esiintyvää metsästyksen jumalatar Juksáhkkáá. Hahmo innosti häntä perehtymään saamelaiseen historiaan ja nykyaikaan.
– Kielitieteilijät arvioivat kalliomaalauksia tehneiden ihmisten puhuneen jotakin pale-oeurooppalaista kieltä, koska saamen- ja suomen kielet olisivat heidän mielestään maalauksia nuorempia, kertoo Honkakoski.
Hän huomauttaa, että saamelaisten historiasta on arkeologista tietoa vasta 2000 vuotta näitä maalauksia myöhemmin. Silti, jos nyt löydetyistä kalliomaalauksista ja jo aiemmin tiedetyistä asioista löydetään yhteneväisyyksiä, ovat tulokset Honkakosken mukaan erittäin tärkeitä.
Suomessa kalliomaalauksia on suhteellisen paljon. Jos tarkastelee laajempaa alu-etta esimerkiksi Norjaan ja Venäjälle saakka, on uusissa löydöksissä yhtäläisyyksiä näistä maista löydettyjen kalliomaalausten kanssa.
Kalliomaalauksista on aiemminkin ilmennyt kuvia, jotka ovat tuttuja saamelaismyto-logiasta: eläimiä, tuntureita, lintuja, ihmisiä metsästystä, kalastusta ja niin edelleen, luettelee Ojanlatva.
Hän lisää,että koko arktisen alueen maalausten tutkiminen olisi mielenkiintoista. Esi- merkiksi kivikautisia maalauksia voitaisiin verrata 200 vuotta vanhoihin saamelais-rumpuihin. Jos jokin taho alkaa tutkimaan kalliomaalauksia,toivoisi Ojanlatva saame- laismuseon voivan olla työssä avuksi. Hän näkee saamelaisalueelta tulevan tiedon tärkeäksi osaksi mahdollista tutkimustyötä.
Arkeologian dosentti: kuvien ajatusmaailma näkyy myös saamelaisrummuissa
Kalliomaalauksista väitöskirjan tehnyt arkeologian dosentti Antti Lahelma on aiem-min tuonut esiin ajatuksen siitä, että vanhojen saamelaisrumpujen kuvamaailmassa on voinut säilyä jotain kalliomaalausajan kulttuurin ajattelutavoista. Pekka Honkakosken kokoama uusi dokumentaatio tukee Lahelman näkemystä.
Lahelman mukaan Honkakosken löydökset ovat mielenkiintoisia: osa on ollut tiedossa jo ennen, mutta osa tulkinnoista on aivan uusia.
– Kuvat ovat yhtäläisiä esimerkiksi Juksáhkkán kanssa, mutta se ei tarkoita sitä, että kuvien tekijät olisivat olleet saamelaisia tai saamelaisuutta olisi edes ollut silloin. Ei ole myöskään tietoa siitä, mitä kieltä he ovat puhuneet, kertoo Lahelma.
Saamelaismuseo Siidan arkeologi Eija Ojanlatva pitää uutta dokumentaatiota kallio-maalauksista tärkeänä ja näkee monenlaisten tutkimusten mahdollisuuden. Vesa Toppari / Yle

– Olivat ne sitte mitä tahansa ihmisiä ja puhuneet mitä tahansa kieltä, on maailman-kuvassa sellaisia ominaisuuksia, joita on myöhemmin nähty saamelaisessa uskossa, hän jatkaa.
Lahelma kuitenkin muistuttaa, että kyse on monia tuhansia vuosia vanhoista maa-lauksista.Vaikka yhteneväisyyksistä saamelaisten kuvausten kanssa puhutaankin, ei voida sanoa, että nämä kalliomaalaukset olisivat saamelaiskulttuurista – toisin kuin vaikkapa rummuissa, joissa yhteys on voitu selvittää.
Lahelma suositteleekin ajattelemaan näitä maalauksia ikään kuin ihmiskunnan yhtei- senä perintönä, eikä niitä kannata viedä hänen mukaansa poliittiseen kontekstiin lainkaan.
https://royalsocietypublishing.org/doi/10.1098/rspb.2019.1528
The genomic ancestry of the Scandinavian Battle Axe Culture people and their relation to the broader Corded Ware horizon
Abstract
The Neolithic period is characterized by major cultural transformations and human migrations,with lasting effects across Europe.To understand the population dynamics in Neolithic Scandinavia and the Baltic Sea area,we investigate the genomes of indi- viduals associated with the Battle Axe Culture (BAC), a Middle Neolithic complex in Scandinavia resembling the continental Corded Ware Culture (CWC).We sequenced 11 individuals (dated to 3330–1665 calibrated before common era (cal BCE)) from modern-day Sweden,Estonia and Poland to 0.26-3.24× coverage.Three of the indivi- duals were from CWC contexts and two from the central-Swedish BAC burial ‘Bergs-graven’. By analysing these genomes together with the previously published data, we show that the BAC represents a group different from other Neolithic populations in Scandinavia,revealing stratification among cultural groups. Similar to continental CWC, the BAC-associated individuals display ancestry from the Pontic–Caspian steppe herders,as well as smaller components originating from hunter-gatherers and Early Neolithic farmers. Thus, the steppe ancestry seen in these Scandinavian BAC individuals can be explained only by migration into Scandinavia. Furthermore, we highlight the reuse of megalithic tombs of the earlier Funnel Beaker Culture (FBC) by people related to BAC. The BAC groups likely mixed with resident middle Neolithic farmers (e.g. FBC) without substantial contributions from Neolithic foragers.
1. Introduction
An influential wave of migration into central Europe from the Pontic–Caspian steppe occurred ca 3000 before common era (BCE) [1,2]. The Yamnaya expansion brought new genetic ancestry to Neolithic Europe, which still today is a part of the genetic variation [3]. Northwest of the Yamnaya complex, we find the Corded Ware Culture (CWC) complex that was distributed over central and northern Europe between 3000 / 2800 and 2300/2000 BCE (see electronic supplementary material) [4,5].The impact of the Yamnaya migration on the formation of the CWC complex continues to be de-bated ([4] with comments in [5,6]). Genetic signals of migration and admixture have been found in central Europe,Scandinavia,and to the east of the Baltic Sea [1,2,7-9]. However, the development took different paths in different regions and it is not clear exactly how the migrations affected the demographic development and population history at the fringes of the European continent [5,6].
In Sweden, the CWC complex has been labelled the (Swedish-Norwegian) Boat Axe or Battle Axe Culture (BAC).The material manifestation of BAC,starting around 3000 / 2800 BCE [10],as distributed in Scandinavia up to modern-day Middle Sweden and southern Norway and on the eastern side of the Baltic Sea up to the southwestern parts of Finland (e.g. [11–13]). How the BAC/CWC complex dispersed into the Baltic countries and Fennoscandia has been extensively debated (e.g. [12,14 – 18]), especially in association with a process of migration or of cultural diffusion and local development [11, 12, 16, 19, 20]. Earlier archaeological research viewed the BAC / CWC complex as a society with common cultural and social practices and stressed the uniformity of burial customs, pottery design and typology, and the boat-shaped battle axes (e.g. [12,15,21]). Recently, it has been argued that these views were too simplistic, and regional patterns and traits within the BAC/CWC complex have been highlighted (e.g. [4–6,11,22]).
While previous archaeogenetic research has studied individuals connected with the BAC /CWC cultural complexes in some regions around the Baltic Sea [1,2,7 – 9,23], the character of the migration patterns and the temporal and regional dynamics within the CWC area as well as the BAC introduction into Scandinavia are yet to be explored. To achieve a better understanding of these population dynamics and the BAC introduction to Scandinavia, we investigated the demographic development in the CWC area around the Baltic Sea, in the third millennium BCE, by sequencing DNA from 11 prehistoric individuals from (modern-day) Sweden,Estonia and Poland. By comparing the genetic profiles of the newly sequenced individuals to individuals from different regions of the CWC area and from different kinds of cultural contexts, we investigate ancestry and admixture to paint a more detailed picture of the demo-graphic processes that took place across the CWC area, with a specific focus on the onset and subsequent dynamics of the BAC in Scandinavia.
2. Results
(a) Genome sequencing, quality assessment, and genetic results
We generated and analysed genome sequence shotgun data from 11 individuals ori-ginating in northeastern Europe dated to 3300–1660 cal BCE (electronic supplemen-tary material,table S1).Five individuals were excavated from CWC contexts:two from Obłaczkowo, Poland, one from Karlova, Estonia and two from the CWC-related BAC burial Bergsgraven in Linköping, Sweden. The six additional individuals were from other archaeological contexts in Sweden:five from megalithic burial structures prima-rily associated with Funnel Beaker Culture (FBC) (two from Rössberga in Västergöt-land and three from Öllsjö in Scania) and one from a Pitted Ware Culture (PWC) context (Ajvide on Gotland). Radiocarbon dating showed that the three individuals from the Öllsjö megalithic tomb derived from later burials, where oll007 (2860 – 2500 cal BCE) overlaps with the time interval of the BAC, and oll009 and oll010 (1930 – 1650 cal BCE) fall within the Scandinavian Late Neolithic and Early Bronze Age (table 1; electronic supplementary material, table S1 and figure S2). Genome-wide sequence coverages range from 0.1 to 3.2×, and the sequence data for all indivi-duals exhibit characteristic properties of ancient DNA: short fragment size and cyto-sine deamination at the ends of fragments (e.g. [24]) (table 1; electronic supplemen-tary material,figure S5). Estimates of mitochondrial contamination [25] were low, less than 2% for all 11 individuals,as was the estimated nuclear contamination on the X-chromosome in males [26,27] (less than 1.2%) (table 1; electronic supplementary material, table S3). Five individuals were genetically determined to be males, and six were females, based on the fraction of sequence fragments mapping to the different sex chromosomes [28] (table 1).
Table 1.
Information on the 11 ancient individuals investigated in this study, including radio-carbon dates,average read length,average genome coverage,average mtDNA cove- rage, mtDNA and Y-chromosome haplogroups, biological sex, and contamination estimates based on mtDNA and on the X-chromosome in males.
The individuals from BAC and CWC contexts, including oll007 from a megalithic bu-rial, displayed U4 and U5 mitochondrial DNA (mtDNA) lineages, previously associa-ted with Stone Age hunter–gatherers [29–34],and H1,N1a and U3 lineages, associa- ted with Neolithic farmers [1,32,35,36] (table 1; electronic supplementary material, table S4). This broadly coincides with the wide variety of mtDNA lineages found in other individuals from CWC contexts (e.g.[2,32]). However,the U3 and N1a lineages, which were found here (poz44 and ber2),have not been reported from individuals ex-cavated in CWC contexts. The two males in our dataset (ber1 and poz81) belonged to Y-chromosome R1a haplogroups (table 1;electronic supplementary material, table S5), as do the majority of males (16/24) from the previously published CWC contexts (Viby in Sweden, Ardu and Kunila in Estonia, Gyvakarai and Spiginas in Lithuania, Bergrheinfeld and Esperstedt in Germany, and Brandýsek in the Czech Republic) [1, 2,7,31,32,37], while a smaller fraction belonged to R1b [3/24] or I2a [3/24] lineages (Tiefbrunn and Esperstedt in Germany,Pikutkowo and Łęki Małe in Poland,and Bran- dýsek in the Czech Republic) [2,23,32,37]. The R1a haplogroup has not been found among Neolithic farmer populations nor in hunter-gatherer groups in central and western Europe, but it has been reported from eastern European hunter-gatherers and Eneolithic groups [1,31,32].Individuals from the Pontic-Caspian steppe, associa- ted with the Yamnaya Culture, carry mostly R1b and not R1a haplotypes [1,2,31,32].
Three individuals had enough data for investigating the LCT gene-region (electronic supplementary material, table S6),and one of these individuals (kar1) carried at least one allele (-13910 C->T) associated with lactose tolerance,while the other two indivi- duals (ber1 and poz81) carried at least one ancestral variant each, consistent with previous observations of low levels of lactose tolerance variants in the Neolithic [1,2, 33,38] and a slight increase among individuals from CWC contexts [32]. The indivi-duals further displayed a mixed appearance with both light and dark hair and brown and blue eyes (electronic supplementary material,table S6). Stable carbon and nitro-gen isotope values for the individuals from modern-day Sweden show a terrestrial diet except for the Ajvide individual (electronic supplementary material, table S1 and figure S3). Strontium isotope data for the two individuals in Bergsgraven have diffe-ring signals, indicating recent migration of at least one of the individuals to the area (electronic supplementary material, table S2 and figure S4).
(b) Exploratory analyses
To investigate the genomic ancestry of the 11 individuals from this study in context with other ancient individuals,we projected them onto the PC1–PC2 space of 60 mo-dern-day Western Eurasian populations (figure 1a) [40] together with 272 relevant prehistoric individuals who have been previously reported (electronic supplementary material, table S7).We further used an unsupervised clustering approach [41] to exa-mine the broad-scale genomic affinities (electronic supplementary material, figure S6).
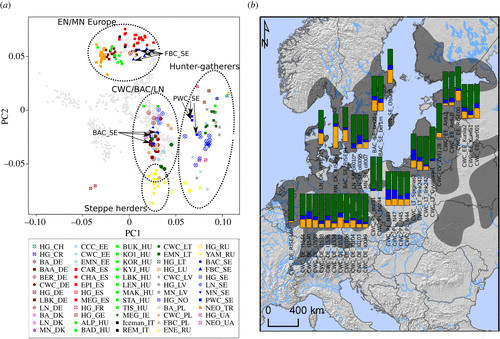
Figure1.(a) Principal component analysis of modern Europeans (grey) and projected ancient Europeans. The full list of ancient samples with sample IDs, the associated culture or time period followed, and two-letter International Organization for Standar- dization codes for country can be found in table 1 and electronic supplementary material,table S7.(b) Geographical range of BAC/CWC and admixture modelling with qpAdm of BAC/CWC individuals in a three-source model:Anatolian Neolithic (orange), European hunter–gatherers (blue), and Yamnaya steppe herders (green). Individuals marked with an asterisk lived after the BAC/CWC time period. Map source: Sjögren et al. [39], published CC-BY.
The broad continental pattern that appears confirms previous results (figure 1a) [3]: (i) a clear separation between Early and Middle Neolithic farmers and Mesolithic hunter-gatherers from all over Europe [1,30,32–34,42,43], (ii) substructure among the hunter–gatherers roughly corresponding to an east–west gradient [1,8,29,31,32], with an exception in Scandinavia [29], and (iii) most Late Neolithic and Bronze Age indivi-duals overlapping with modern-day central and northern Europeans due to ad-mixture with incoming groups related to the Yamnaya herders from the Pontic steppe [1,32].
In Scandinavia, we see a clear separation between individuals from the three diffe-rent archaeological complexes, the FBC, PWC,and BAC, with individuals associated with the farming FBC (including the Rössberga individuals reported here) grouping with other Middle Neolithic farmer groups (figure 1a). Neolithic foragers associated with the Pitted Ware Culture (including ajv54) were genetically similar to Mesolithic Scandinavian hunter–gatherers with some trend towards farming groups, likely due to admixture between the two groups [9,29,33]. The individuals associated with BAC (including the two individuals from Bergsgraven) show a clear association with indivi-duals from other CWC contexts elsewhere in Europe. Notably, the oll007 individual from the megalithic site at Öllsjö (southern Sweden), who was not directly associa-ted with any CWC-related artefacts but overlaps with CWC chronologically, clusters with individuals from CWC contexts, as do the two individuals with later dates (oll009 and oll010) from the same site.
Similar to the BAC individuals,the newly sequenced individuals from the present-day Karlova in Estonia and Obłaczkowo in Poland appear to have strong genetic affini-ties to other individuals from BAC and CWC contexts across the Baltic Sea region [1, 2, 7, 8, 23,31,32] (figure 1a). Some individuals from CWC contexts, including the two from Obłaczkowo, cluster closely with the potential source population of steppe-related ancestry, the Yamnaya herders.Notably,these individuals appear to be those with the earliest radiocarbon dates among all genetically investigated individuals from CWC contexts. Overall, for CWC-associated individuals, there is a clear trend of decreasing affinity to Yamnaya herders with time (figure 2).
{kind=link}

Figure 2. Correlation between f4(Chimp, LBK, YAM, X), where X is a CWC or BAC individual, and the date (BCE) of each individual (table 1 and [1,2,7,8,23,31, 32, 37]). This statistic measures shared drift between CWC and Linear Pottery Culture (LBK) as opposed to YAM and should increase with the higher proportion of Neolithic farmer ancestry in CWC and BAC. The pattern is not driven by spurious affinities between single nucleotide polymorphism (SNP) capture or shotgun data (electronic supplementary material, figures S9 and S10). Ages on the x-axis correspond to the mid-point of the interval for the date of each sample as reported in their original publication (electronic supplementary material, tables S7 and S8). (Online version in colour.)
{kind=link}
(c) Admixture modelling of Corded Ware Culture
To investigate the spread of CWC-related people into Scandinavia and obtain a clea-rer picture of the ancestry proportions found in individuals associated with the CWC and the BAC,we conducted a supervised modelling of their ancestry from three sour-ces: Anatolian farmers, western European hunter–gatherers, and Yamnaya steppe herders. We estimated ancestry under this three-source model per individual using qpAdm [1]. The geographical pattern of ancestry is shown in figure 1b.This approach confirms that most ancestry in all CWC individuals originates from a group related to Yamnaya steppe herders (green), while smaller contributions from Western hunter–gatherers (blue) and Neolithic farmers (orange) are also widespread (figure 1b).
Individuals from BAC contexts in (modern-day) Sweden also show this pattern, with a large genetic component tracing its ancestry to Yamnaya steppe herders and with small ancestry components related to Neolithic farmers and hunter–gatherers. The other individuals who were contemporary with BAC but had unclear cultural contexts and who were buried in the Ölljsö megalith constructed many hundred years earlier (oll007), or found as a stray find (Ölsund) [9],show the same genetic profile as indivi-duals from typical BAC contexts in other parts of Sweden. Although the individuals associated with the BAC (including the oll007, oll009, oll010, and Olsund) have a large proportion of steppe ancestry, it is relatively low compared with most other indi-viduals from CWC contexts. The female (kar1) from the Karlova CWC burial in Esto-nia shows similar patterns, also displaying slightly less steppe ancestry than other CWC-associated individuals from Estonian sites [2,7]. By contrast, the CWC indivi-duals from Obłaczkowo in Poland (poz44 and poz81) show an extremely high pro-portion of steppe ancestry (greater than 90%), which is different from the later CWC-associated individuals excavated in Pikutkowo (Poland) [23], but similar to some other CWC-associated individuals from Germany, Lithuania, and Latvia [2,8,31].
Interestingly, these individuals with a large fraction of steppe ancestry have typically been dated to more than 2600 BCE, making them among the earliest CWC indivi-duals genetically investigated.This observation, i.e.early CWC individuals resembled (genetically) Yamnaya-associated individuals, while later CWC groups show higher levels of European Neolithic farmer ancestry (Pearson's correlation coefficient: −0.51, p = 0.006) (figure 2), suggests an initial dispersal that occurred rapidly.
(d) Admixture modelling of Battle Axe Culture
The positioning in the principal component analysis (PCA) (figure 1a) and the admix-ture modelling results (figure 1b) suggest ancestry from three different main ancest-ral groups in BAC: Anatolian farmers, European hunter–gatherers, and Yamnaya steppe herders.Direct candidates for contributions to BAC are the Scandinavian Middle Neolithic PWC and FBC plus a third population carrying high proportions of Yamnaya-related ancestry (YAM),like CWC. We used an explicit model to investigate contri-butions from predefined ancestral groups (qpWave of ADMIXTOOLS) [1,44] to model the BAC genetic make-up. We found that the BAC-associated individuals can be modelled as a combination of genetic material from these three groups (p= 0.80). In ad-dition to the three-source model, two models only using two sources were con-sistent with the data: the best fitting two-source model was FBC+ YAM (p = 0.86), while YAM + PWC would also fit the data (although trending toward low p-values;p = 0.07). These observations suggest that - to our statistical resolution - a direct PWC contri-bution to BAC is not needed in a model, but actual PWC admixture might have been small or there may have been indirect PWC contributions through PWC first mixing with FBC [34] who later contributed ancestry to BAC. Notably, using only the CWC population as the single source for BAC was consistent with the data in all ca-ses (p >0.05,except when using CWC-associated individuals from Latvia, CWC_LV), showing that the BAC individuals in Sweden could not have emerged without migrations from other CWC groups.
To find out if BAC represents an admixed group that came into Scandinavia or if (at least some of) the admixture took place in Scandinavia, we investigated the relation-ship among the Scandinavian BAC group and various CWC groups from other geo-graphical areas by constructing admixture graphs (figure 3). The BAC groups fit as a sister group to the CWC-associated group from Estonia (CWC_EE,electronic sup-plementary material,figure S8) but not as a sister group to the CWC groups from Po- land (CWC_PL, figure 3) or Lithuania (CWC_LT, electronic supplementary material) (|Z|>3),indicating some differences in ancestry between these CWC groups and BAC (CWC from Latvia, CWC_LV, and the CWC group from Germany, CWC_DE, had to be excluded from this analysis due to the number of SNPs being too low, see elect-ronic supplementary material). Supervised admixture modelling suggests that BAC may be the CWC-related group with the lowest YAM-related ancestry and with more ancestry from European Neolithic groups (figure 1b). Consequently, models, where BAC is an admixed group between a CWC group and additional ancestry from a group carrying ancestry-related European Neolithic groups, were consistent with the data for all three tested CWC groups (|Z| < 2.3, figure 3; electronic supplementary material, figures S7 and S8).
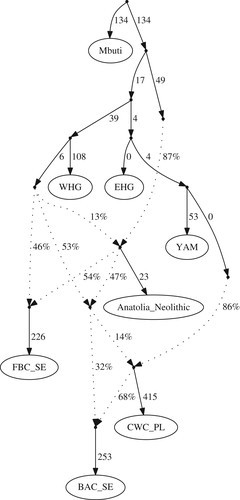
Figure 3.Admixture graph modelling BAC as a mix of a CWC source and an admixed Middle Neolithic group (worst |Z| < 1.68). CWC individuals from modern-day Poland (CWC_PL) chosen here, other groups shown in electronic supplementary material, figures S7 and S8. WHG, western hunter-gatherers; EHG, eastern hunter-gatherers. The sub-Saharan African Mbuti were used as an outgroup for this analysis.
{kind=link}
Two potential scenarios were consistent with the data to explain the emergence of BAC in Scandinavia: (i) direct migration of CWC groups from the eastern Baltic or (ii) a migration of CWC groups from the southern Baltic Sea region mixing with FBC groups in Scandinavia. Future archaeogenetic studies could fill geographical and chronological gaps by including more samples that should help to distinguish between the scenarios.
3. Discussion
The gene pool of modern Europeans has been shaped by a number of prehistoric events and migrations. The CWC horizon represents one of the major demographic processes in the northern areas of Europe as this is associated with the first occurrence of steppe-related ancestry. Our results have implications for our view on the demographic development associated with the CWC in general, and the Scandinavian variety of the BAC specifically.
People buried in CWC contexts display a genetic ancestry component that was not present in northern and central Europe prior to the third millennium BCE.This ances-try component, often called ‘steppe ancestry’, probably traces back to the ‘Yamnaya expansion’ of herders associated with the Yamnaya Culture that dispersed into Eas-tern Europe from the Pontic-Caspian steppe around 3000 BCE [1,2,32]. This compo-nent makes up the largest proportion of the genetic ancestry in all sequenced BAC / CWC individuals around the Baltic Sea: from the modern-day countries Estonia, Lat-via, and Lithuania in the east; Poland and Germany in the southwest; and Denmark and Sweden in the northwest (figure 1b). One important observation is that the ear-liest CWC individuals analysed to date had the highest proportion of steppe ancestry (greater than 90%), while this proportion decreased in later individuals (figure 2).
This suggests a gradual process of admixture between incoming groups and local groups, such as the FBC groups in northern Europe that traced most of their genetic ancestry to Anatolian Neolithic farmers. This process was driven by in-coming males mixing mainly with local females [5,7,45,46].The admixture process is evident across the en-tire distribution of the CWC, even in regions such as the eastern Baltic coast where no FBC groups or genetically related groups have been found. Consequently, farmer-related ancestry must have arrived in the eastern Baltic region via migrations more recent than 2500 BCE [7–9].Possibilities include a gene-rally large exchange network across the entire CWC horizon or specific migrations into the eastern Baltic region. Potential source regions for the latter could be modern-day Poland or Sweden, where FBC groups predating the arrival of the CWC are found.
The paternal lineages found in the BAC/CWC individuals remain enigmatic. The ma-jority of individuals from CWC contexts that have been genetically investigated this far for the Y-chromosome belong to Y-haplogroup R1a, while the majority of sequen-ced individuals of the presumed source population of Yamnaya steppe herders be-long to R1b [1,2]. R1a has been found in Mesolithic and Neolithic Ukraine [1,31,32]. This opens the possibility that the Yamnaya and CWC complexes may have been structured in terms of paternal lineages—possibly due to patrilineal inheritance sys-tems in the societies [47,48]-and that genetic studies have not yet targeted the direct sources of the expansions into central and northern Europe.
The Scandinavian Middle Neolithic megalithic tombs are associated with the FBC. However, their reuse, indicated by artefacts common to the BAC and later periods, has been noted [17].The oll007 individual,buried in the FBC-associated Öllsjö mega- lithic tomb, but radiocarbon dated to the time period of the BAC, is genetically very similar to individuals from BAC contexts (e.g. Bergsgraven and Viby). Thus, although archaeologically the reuse of megalithic tombs was assumed earlier [17], our study may be the first direct link (using genetics) showing that indeed FBC-associated me-galithic tombs were used as burial places also for the people of the BAC. This could possibly also extend to the Danish Single Grave Culture (SGC) [49], as RISE61 [2], a male buried in the Kyndeløse passage grave and with a radiocarbon date overlap-ping with the BAC/CWC/SGC time period, also displays some steppe ancestry.
The BAC replaces the FBC in the southern parts of Scandinavia and was previously assumed to have been a cultural adaptation of existing groups [12,17]. We show in multiple individuals from different parts of Scandinavia that these groups (BAC) were part of the general CWC horizon, i.e. they too are the result of admixture of different groups tracing parts of their ancestry to European hunter–gatherers, Anatolian Neoli-thic farmers, and Yamnaya steppe herders. This implies that BAC groups were not the direct descendants of any of the groups that lived in the area previously or even contemporaneously—i.e. the groups associated with FBC or the PWC. We also note that the BAC group does not have a particular genetic connection with other eastern Baltic groups such as the Combed Ceramic Culture. The mixed ancestry of indivi-duals in BAC contexts is evident across all autosomal analyses, as well as in mito-chondrial haplogroups,but the paternal haplogroups stand out to some extent, show-ing a deviant,more extreme,pattern. This Y-chromosome pattern is, however, consis- tent with a male sex-biased migration and admixture process among the Yamnaya, and later CWC, groups [7,39,45,50].
As the individuals of the BAC complex cannot be modelled as direct genetic descen-dants of FBC or PWC groups, a migration into Scandinavia of people with a large proportion of steppe ancestry must have taken place.We were not able to find unam-biguous evidence for a specific source population by testing all other individuals as-sociated with the CWC that have been genomically investigated as potential ances-tors.The Scandinavian BAC group has more Neolithic farmer ancestry than pre-2600 BCE individuals in CWC contexts from the southern or eastern Baltic coast, sugges-ting that they mixed with an FBC group. This process could have happened in Scan-dinavia or before arriving in Scandinavia. On the one hand, the CWC people were mainly bound to travels on land, which would favour migration from central Europe into modern-day Denmark and Sweden (e.g. [16]). On the other hand, there is evi-dence for technological exchange (pottery craft) crossing the Baltic Sea in this time period [20,51],which may or may not be associated with gene flow. Finally, the gene-tic data from BAC contexts are still limited, and the patterns of gene flow that we ob-serve are consistent with both a single migration event into Scandinavia and with a continuous process with an extensive network of social and technological exchange. Future studies will refine our understanding of the social, geographical, and temporal dynamics during this important period in European prehistory.
4. Material and methods
We extracted DNA, prepared Illumina DNA libraries [29,33,34], and shotgun sequen-ced DNA from 11 archaeological remains from Sweden, Estonia,and Poland (table 1; electronic supplementary material). The samples were radiocarbon dated (n = 10) and analysed for stable isotope values (carbon,n = 8;nitrogen, n = 8; strontium, n = 2). Sequence data were merged, trimmed, and mapped to the human genome, and the data were authenticated using fragmentation, deamination patterns, and various con-tamination estimates [29]. Mitochondrial and Y-chromosomal DNA haplogroups were called for the newly sequenced individuals. The comparative data consisted of an-cient individuals with at least 0.1×genome coverage who lived at the time of the CWC or before (electronic supplementary material, table S7) [1,2,7–9,23, 29 – 34, 42, 43, 52 – 59] and modern populations from the Human Origins dataset [30] and the public data of the Simons Genome Diversity Panel [60]. Ancestry and admixture were assessed using principal component analyses, unsupervised ADMIXTURE [41], and admixture proportions were estimated using ADMIXTOOLS' qpWave and qpAdm [1, 44], and phenotypic variation was investigated [61,62]. More detailed descriptions of the archaeological samples and sites and the methods used can be found in the electronic supplementary material.
Data accessibility
The genome data for the 11 investigated individuals are available at the European Nucleotide Archive (accession number PRJEB32786).
Authors' contributions
H.M., T.G., A.G., J.S., and M.J. designed the research; H.M.,T.G., E.M.S., A.J., M.F., A.R.M., A.G., J.S., and M.J. performed the research; H.M., A.J., M.F., Ł.P., M.T., J.L., and J.S. contributed samples and conducted archaeological analyses; H.M., T.G., and E.M.S. analysed data; and H.M., T.G., A.G., J.S., and M.J. wrote the paper with input from all authors.
Competing interests
We declare we have no competing interests.
Funding
This work was supported by the Berit Wallenberg Foundation (grant no. BWS2011.0090 (M.F.)); the Swedish Research Council (grant nos 2017-02503 (H.M.), 2017-05267 (T.G.), 2013-1905 (M.J.,J.S.,and A.G.));Riksbankens Jubileums- fond (grant no. M13-0904:1 (M.J., J.S., and A.G.)) and Knut and Alice Wallenberg foundation (M.J., J.S., and A.G.). The Ministry of Science and Higher Education of the Republic of Poland funded the radiocarbon dates for poz44 and poz81 (grant no. N N109 287 137 (L.P.)).
Acknowledgements
We thank the following museums and persons for supplying bone material: P. Nils-son, M. Ohlsén, P. Nyberg, and M. Douglas from Östergötlands Museum, L. Drenzel and J. Karlsson at the National Historical Museums (Stockholm), the Historical Museum at Lund University (LUHM), P. Pawlak from Henryk Klunder's Pracownia Archeologiczno-Konserwatorska (Poznań) and Ü. Tamla and the Archaeological Research Collections, Tallinn University, Estonia. Further, we thank J. Evans at NERC, Isotope Geosciences Laboratory, Nottingham, UK, for performing strontium isotope analyses and T. Hedlund for osteological analyses on the Öllsjö individuals. Shotgun sequencing was performed at the National Genomics Infrastructure (NGI) Uppsala, and computations were performed at Uppsala Multidisciplinary Center for Advanced Computational Science (UPPMAX) under projects b2013203, b2013240, b2015066, snic2017-7-259, sllstore2017020, and sllstore2017087.
Footnotes
†These authors contributed equally to this study.
Electronic supplementary material is available online at https://doi.org/10.6084/m9.figshare.c.4682318.
Published by the Royal Society under the terms of the Creative Commons Attribution License http://creativecommons.org/licenses/by/4.0/, which permits unrestricted use, provided the original author and source are credited.
***
Uuutta humpuukia:
https://www.friatider.se/studie-skandinaviens-forsta-befolkning-utrotades-av-invandrare

Studie: Skandinaviens första befolkning utrotades av invandrare
Publicerad 5 februari 2024 kl 15.40Inrikes. När de första bönderna kom till Skandinavien för 5.900 år sedan utrotades samlar- och jägarbefolkningen inom ett par generationer. Det visar en ny studie som publiceras i Nature. Resultaten – som går på tvärs mot rådande uppfattning – bygger på dna-analyser av skelett och tänder som hittats i dagens Danmark.
Dela artikeln
Det internationella forskarlaget har genom att extrahera dna från skelettdelar och tänder från forntida människor kunnat dra nya slutsatser om migrationens effekter på förhistoriska populationer.
Studien visar bland annat att det skett två nästan totala populationsförändringar i Danmark under de senaste 7.300 åren. Det första befolkningsbytet skedde för 5.900 år sedan då en bondebefolkning, av olika härkomst och utseende, fördrev de samlare, jägare och fiskare som tidigare befolkat Skandinavien.
Inom loppet av bara några generationer utrotades nästan hela jägar- och samlarbefolkningen, enligt forskarna.
– Det har tidigare framställts som om denna övergång varit fredlig, bland annat i tv-serien "Historien om Danmark" och motsvarande serie i Sverige. Men vår studie tyder på det motsatta. Förutom ond bråd död är det troligt att nya patogener från boskap tog kål på många samlare, säger Anne Birgitte Nielsen, geologiforskare och föreståndare för Laboratoriet för 14C-datering vid Lunds universitet.
Ett tusental år senare, för cirka 4.850 år sedan, skedde ytterligare en befolkningsförändring när människor med genetiska rötter i Yamnaya – ett indoeuropeiskt herdefolk med rötter i södra Ryssland – tog sig till Skandinavien och utrotade den tidigare bondebefolkningen. Återigen kan det enligt forskarna både ha hänt med våld och genom nya patogener. De storvuxna människorna förde ett halvnomaniserade liv på stäpperna, tämjde djur, höll tamboskap och flyttade sig med häst och vagn över stora områden. De människor som slog sig ned på våra breddgrader var en blandning mellan Yamnaya och östeuropeiska bondestenåldersmänniskor – en genetisk profil som än idag är dominerande i Danmark medan den första bondebefolkningens dna-profil i princip är raderad.
– Även denna gång skedde befolkningsförändringen snabbt, och praktiskt taget utan ättlingar från föregångarna. Vi har inte ett lika stort dna-material från Sverige, men det som finns pekar på ett liknande händelseförlopp. Många svenskar härstammar med andra ord också i stor utsträckning från dessa halvnomader, säger Anne Birgitte Nielsen i ett pressutskick.
Resultaten omkullkastar inte bara tidigare teorier om amorösa och fredliga möten mellan folkgrupper. Studien ger även en fördjupad förståelse för de historiska migrationsströmmarna samt viktiga nycklar till hur man kan tolka arkeologiska fynd och förändringar i vegetation och arealanvändning som påträffats i paleoekologiska data.
– Våra resultat bidrar till en djupare kunskap om vårt arv. Men också förståelse för utvecklingen av vissa sjukdomar. Något som på längre sikt kan bli nyttig i exempelvis medicinsk forskning, säger Anne Birgitte Nielsen. "
Kommentit